Los brasinoesteroides. Una nueva clase de hormonas vegetales
Introducción
Las hormonas vegetales son sustancias fabricadas en un determinado lugar de una planta y que se transladan a otro donde actúan a muy bajas concentraciones, regulando el crecimiento, desarrollo o metabolismo del vegetal. Los cambios en la concentración de la hormona y la sensibilidad de los tejidos provocan una amplia gama de efectos en las plantas, muchos de los cuales involucra interacciones con el ambiente como un tipo de adaptación, considerando que las plantas son organismos inmóviles. Para diferenciarlos de las hormonas vegetales de origen natural, los compuestos que producen efectos fisiológicos pero que son de origen sintético se denominan “reguladores del crecimiento vegetal”.
Se reconocen comúnmente como hormonas vegetales cinco tipos de compuestos: auxinas, citocininas, giberelinas, etileno y ácido abscísico. Cada uno de éstos tiene efectos fisiológicos propios. Por ejemplo, las auxinas están involucradas en el alargamiento celular. Las citocininas inducen la división celular, retardan el envejecimiento (senescencia) de las hojas, etcétera. Las giberelinas inducen la germinación de semillas y la floración, etcétera. El ácido abscísico induce la latencia de las semillas, el cierre de los estomas (estructuras respiratorias de la planta), etcétera. El etileno, a pesar de ser un compuesto muy simple químicamente, es capaz de inducir la maduración de ciertos frutos.
Además de estas cinco fitohormonas, consideradas clásicas, en la década pasada se obtuvieron fuertes evidencias científicas de que existen otros compuestos involucrados en diferentes procesos del desarrollo de las plantas, como las poliaminas, el ácido jasmónico, el ácido salicílico y los brasinoesteroides. De estos últimos hablaremos aquí ampliamente.
Descubrimiento de los brasinoesteroides
A principios de la década de los sesenta, algunos investigadores tenían la hipótesis de que la germinación acelerada y el crecimiento de los granos de polen podrían estar asociados con la presencia de promotores del crecimiento. En 1970 Mitchell y colaboradores reportaron que algunos extractos del polen de colza (Brassica napus L.) producían un poderoso efecto de elongación (alargamiento) en el tallo de frijol. Esta respuesta fue distinta a la que producen otras hormonas denominadas giberelinas. Las sustancias que promovieron el crecimiento de manera más activa fueron aisladas de Brassica napus, y por ello fueron llamados “brassinos”. Mitchell y colaboradores, de manera profética, atribuyeron el estatus de hormona vegetal a los brassinos porque eran compuestos orgánicos específicos, aislados de plantas y que habían inducido crecimiento cuando eran aplicados en cantidades diminutas a otras plantas. Sin embargo, pronosticaron incorrectamente que su estructura era similar a la de los ácidos grasos.

La verdadera naturaleza química de los “brassinos” no fue descubierta sino hasta después de un gran esfuerzo coordinado en diferentes laboratorios de Estados Unidos. En una planta piloto se extrajo una carga de 227 kilogramos de polen de B. napus, dando como resultado cuatro miligramos de una sustancia que fue identificada por cristalografía de rayos X como una lactona esteroidal, y fue denominada brasinólido. Dos años después el brasinólido y su isómero (variante química), el 24-epi-brasinólido, habían sido sintetizados químicamente, eliminando la necesidad de una obtención masiva de extractos de plantas. Con una amplia variedad de compuestos sintéticos a la mano, en la década de 1980 muchos investigadores se enfocaron a determinar sus efectos fisiológicos en una gran diversidad de sistemas biológicos.
 |
El término de brasinoesteroides fue asignado por Mandava (1988) a los esteroides que promovían el crecimiento vegetal en el bioensayo de elongación del segundo internado del frijol. En los inicios de 1990 varios grupos japoneses mostraron avances prometedores en el descubrimiento de la ruta de biosíntesis de los brasinoesteroides, mientras diversos grupos de químicos y biólogos se involucraron en la investigación para determinar si estos compuestos eran una nueva clase de hormonas vegetales. Sin embargo, no fue hasta 1996 cuando se encontraron pruebas contundentes de que eran indispensables para el crecimiento y desarrollo de las plantas: los reportes de cuatro trabajos independientes donde se mostraba la identificación y características de una mutante insensible a los brasinoesteroides y tres mutantes deficientes (es decir, que no contenían ciertos brasinoesteroides o los contenían en cantidades menores a las normales). Las mutantes presentaban enanismo y otras alteraciones fisiológicas que pudieron ser revertidas con la adición de los brasinoesteroides. Aún más, se demostró que dos de las mutantes deficientes resultaron tener alteraciones en los genes de las proteínas involucradas en la síntesis de estos compuestos. De esta manera se tuvo evidencia tanto genética como bioquímica de que los brasinoesteroides estaban involucrados en el crecimiento y desarrollo normal de las plantas. Los brasinoesteroides fueron entonces aceptados por la mayoría de los científicos como una nueva clase de hormonas vegetales, y actualmente el número de investigadores que estudian sus efectos se ha incrementado considerablemente.
Existen evidencias de que estos compuestos, al igual que las giberelinas y las auxinas, están ampliamente distribuidos en el reino vegetal, tanto en plantas superiores como inferiores. Los brasinoesteroides se han encontrado principalmente en polen, hojas, yemas, flores y semillas en proporciones y formas diferentes.
Biosíntesis de los brasinoesteroides
Las moléculas de los brasinoesteroides cuentan con cuatro anillos (A, B, C y D) y una cadena lateral (Figura 1). Son formados a partir de la condensación de bloques de cinco átomos de carbonos denominados isoprenos. Los brasinoesteroides son esteroides con 27, 28 o 29 átomos de carbono con diferentes substituyentes en los anillos A y B y en la cadena lateral. Químicamente se han identificado más de 50 brasinoesteroides de fuentes vegetales. El brasinólido es hasta ahora el que produce la mayor actividad biológica entre ellos, y puede sintetizarse directamente del campesterol o a través de la síntesis general de los esteroles. Los esteroles vegetales, además de su papel como precursores de los brasinoesteroides, son componentes integrales de las membranas celulares, donde regulan su fluidez y permeabilidad.
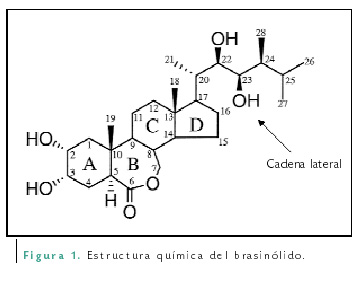 Como se mencionó anteriormente, el término “brasinoesteroides” fue asignado a los esteroides que promueven el crecimiento de los tejidos vegetales. Sin embargo, existen varios esteroides intermediarios en su síntesis, por lo que existía la duda de cuál era la característica de los brasinoesteroides. Para clarificar esta situación, Bishop y Yokota propusieron definir a los brasinoesteroides como esteroides que tengan un oxígeno en el átomo de carbono C-3 y otros adicionales en C-2, C-6, C-22 y C-23 (de acuerdo al orden numérico de los carbonos de los esteroides, Figura 1).
Como se mencionó anteriormente, el término “brasinoesteroides” fue asignado a los esteroides que promueven el crecimiento de los tejidos vegetales. Sin embargo, existen varios esteroides intermediarios en su síntesis, por lo que existía la duda de cuál era la característica de los brasinoesteroides. Para clarificar esta situación, Bishop y Yokota propusieron definir a los brasinoesteroides como esteroides que tengan un oxígeno en el átomo de carbono C-3 y otros adicionales en C-2, C-6, C-22 y C-23 (de acuerdo al orden numérico de los carbonos de los esteroides, Figura 1).
Efectos fisiológicos
La aplicación de los brasinoesteroides induce un amplio rango de respuestas, incluyendo un incremento en la tasa de elongación del tallo, aumento en la expansión de las hojas, crecimiento del tubo polínico, desenrollamiento de las hojas en pastos, reorientación de las microfibrillas de celulosa, así como la adaptación al estrés, ya que aumenta la tolerancia al frío en plantas de arroz. Un efecto interesante se presenta en células de zinnia (una planta ornamental), donde la adición de los brasinoesteroides induce la formación de tejido conductor.
Las plántulas recién germinadas en la oscuridad, proceso conocido como etiolación, es un ejemplo que muestra cómo la ausencia de luz afecta la morfología de las plantas (fotomorfogénesis). Las plántulas germinadas y crecidas en la oscuridad presentan tallos alargados, y la parte final del tallo donde comienzan las hojas primarias (cotiledones) está curvado en forma de gancho. Además, contienen precursores no diferenciados de los cloroplastos, por lo que el color de las hojas es amarillo pálido. Sin embargo, cuando estas plántulas se exponen a la luz, la elongación del tallo disminuye dramáticamente, la plántula se endereza y aparecen las primeras hojas verdes. Algunos tipos de mutantes con defectos en la síntesis o percepción de brasinoesteroides muestran una morfología similar a las plántulas que crecen en luz aun cuando crecen en la oscuridad. Además, muestran defectos en su desarrollo cuando son cultivados en luz: son enanas, con hojas verdes, con tallos, raíces, pecíolos, hojas y tallos de inflorescencias más cortos que las plantas normales. Asimismo, la fertilidad masculina se ve reducida. De esta manera se deduce que los brasinoesteroides están involucrados directa o indirectamente en la fotomorfogénesis.
Por otro lado, los brasinoesteroides también tienen efecto en los procesos de propagación in vitro o micropropagación. Esta última es un método alternativo para la multiplicación masiva de especies vegetales. Se realiza por medio del cultivo de células vegetales, tejidos o aislamiento de órganos de una planta madre en un medio nutritivo artificial bajo condiciones estériles. Los métodos usados para micropropagar especies se basan en dos procesos, principalmente: la organogénesis, que consiste en la formación de nuevos brotes a partir de diferentes tejidos, y la embriogénesis somática, que consiste en la formación de embriones a partir de células no sexuales como las de hojas, tallos, raíces, etcétera. Actualmente los brasinoesteroides están siendo investigados cada vez más durante el proceso de micropropagación, ya que se ha reportado que estimulan la formación de nuevos brotes, la regeneración de plántulas a partir de embriones somáticos, y recientemente se ha determinado que promueven la formación de los embriones somáticos.

Modo de acción
La expansión celular es un requerimiento crítico para el crecimiento y la formación de órganos en los tejidos vegetales. Está controlado por la alteración coordinada de las propiedades mecánicas de la pared celular, los procesos bioquímicos y la expresión de genes. La pared primaria en la mayoría de las plantas consiste en microfibrillas de celulosa (polímeros de un tipo de azúcar) unidas en una red a la hemicelulosa, los cuales, a su vez, están embebidos en una matriz gelatinosa de pectinas. Para que la expansión celular ocurra, la pared celular debe relajarse brevemente o romper las interacciones con la hemicelulosa, acompañada por la adición de nuevos componentes de la pared celular para prevenir el adelgazamiento y rompimiento de las paredes. La regulación de las síntesis y actividad de las proteínas que modifican la pared celular, como la endotransferasa de xiloglucano, glucanasas, expansinas y celulosa sintasa, se convierten en un blanco obvio de las hormonas involucradas en la elongación celular. Para apoyar este modelo, se ha visto que la adición exógena de los brasinoesteroides en explantes vegetales induce la expresión de diferentes genes, como la endotransferasa de xilanoglucano o las expansinas.
Por otro lado, la dirección de la expansión celular está regulada por la orientación de las microfibrillas de celulosa de la pared celular. Ésta a su vez es controlada por la orientación de microtúbulos corticales. Existen evidencias de que los brasinoesteroides promueven la orientación de los microtúbulos, y de esta manera se acelera la elongación celular.
La evidencia reciente también muestra que los brasinoesteroides estimulan la producción de una cinasa dependiente de ciclina y de la ciclina D; ambas enzimas forman parte de un complejo proteico que activa la división celular. De esta manera se sugiere que éste es otro mecanismo por el que los brasinoesteroides inducen el crecimiento de los tejidos vegetales.
En cuanto al efecto fotomorfogénico característico de las mutantes con defectos en la síntesis o percepción de los brasinoesteroides, existe evidencia que muestra que es posible que la cascada de señales que percibe la luz active a una proteína denominada citocromo P450, proteína clave en la síntesis de los esteroides vegetales. De esta manera existe una relación entre la luz y los brasinoesteroides.
Por otro lado, como se mencionó anteriormente, en general las hormonas vegetales no actúan solas, sino en conjunto. De esta manera se ha determinado que la adición de brasinoesteroides estimula la producción de etileno. Sin embargo, la hormona vegetal con la que parecen tener más relación son las auxinas, principalmente en los efectos fisiológicos de crecimiento, elongación, crecimiento de raíces, etcétera. Se ha demostrado que los brasinoesteroides activan la expresión de genes que normalmente se expresan solamente durante la adición de auxinas. Estas evidencias demuestran que los brasinoesteroides actúan directamente o en conjunto con las auxinas; además, pueden estar sensibilizando a las células de alguna manera a la acción de éstas.
Percepción de los brasinólidos por la célula
 Para que un compuesto ejerza un efecto fisiológico, es necesario que la célula blanco lo perciba. Esta percepción del compuesto puede ser a través de un receptor localizado en la membrana que rodea a la célula, como ocurre con las hormonas en animales. En el caso de los brasinoesteroides, la primera evidencia de que existía un receptor surgió de una planta mutante que no mostraba cambios fisiológicos con la adición de los brasinoesteroides. Pronto se presentaron más pruebas de que esta mutante tenía un defecto en su receptor. Para comprobarlo, Joanne Chory y colaboradores construyeron un gen que tenía el código para la producción de un fragmento extracelular del receptor denominado BRI1, fusionado a una proteína (Xa21) que produce una respuesta letal en presencia de un patógeno. El gen de estas proteínas fusionadas se introdujo en células de arroz con técnicas especiales. La aplicación del brasinólido a estas células produjo su muerte, una respuesta similara la observada cuando las células que expresan el gen Xa21 son expuestas al patógeno. Las células que no contenían la proteína no sufrieron alteración alguna. Este experimento demostró de manera contundente que la proteína BRI1 era el receptor de los brasinoesteroides.
Para que un compuesto ejerza un efecto fisiológico, es necesario que la célula blanco lo perciba. Esta percepción del compuesto puede ser a través de un receptor localizado en la membrana que rodea a la célula, como ocurre con las hormonas en animales. En el caso de los brasinoesteroides, la primera evidencia de que existía un receptor surgió de una planta mutante que no mostraba cambios fisiológicos con la adición de los brasinoesteroides. Pronto se presentaron más pruebas de que esta mutante tenía un defecto en su receptor. Para comprobarlo, Joanne Chory y colaboradores construyeron un gen que tenía el código para la producción de un fragmento extracelular del receptor denominado BRI1, fusionado a una proteína (Xa21) que produce una respuesta letal en presencia de un patógeno. El gen de estas proteínas fusionadas se introdujo en células de arroz con técnicas especiales. La aplicación del brasinólido a estas células produjo su muerte, una respuesta similara la observada cuando las células que expresan el gen Xa21 son expuestas al patógeno. Las células que no contenían la proteína no sufrieron alteración alguna. Este experimento demostró de manera contundente que la proteína BRI1 era el receptor de los brasinoesteroides.
 |
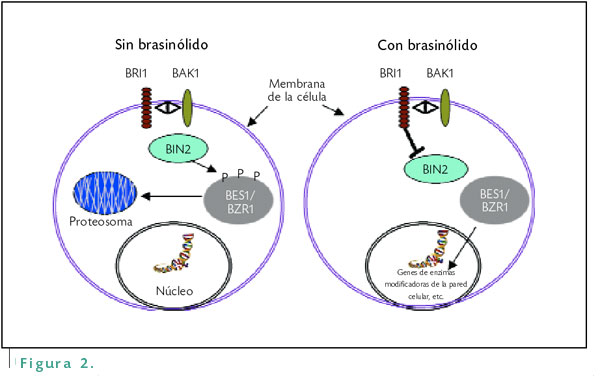
Por otro lado, para ejercer su función el receptor debe interactuar con otros componentes proteicos. Tal es el caso de un componente denominado BIN2, el cual en ausencia del brasinólido le agrega un grupo fosfato a otros componentes conocidos como BES/BRZ1, los cuales al ser fosforilados son degradados por un complejo proteosómico. Por otra parte, si hay presencia del brasinólido, el componente BIN2 no actúa: los componentes BES1/BRZ1 no se fosforilan y por lo tanto no se degradan, y de esta manera estos componentes se acumulan en el núcleo induciendo la expresión de ciertos genes descritos anteriormente, cuyo resultado final es producir una respuesta fisiológica específica (Figura 2).
 |
|
En ausencia del brasinólido, la proteína BIN2 fosforila (P) al complejo BES1/BZR1 de manera que se desestabiliza y es degradado por un complejo proteosómico. Por el contrario, cuando el receptor BRI1 percibe al brasinólido, éste inactiva a la proteína BIN2 y el complejo BES1/BZR1 se acumula en el núcleo, activando la expresión de diferentes genes, incluyendo a los involucrados en la modificación de la pared celular y el crecimiento. La proteína BAK1 interactúa con el receptor BRI1; sin embargo, no se sabe cuál es su función durante la percepción del brasinólido.
|
Perspectivas
En general, en los últimos años se ha generado bastante información sobre la ruta de biosíntesis de los brasinoesteroides y sobre su modo de percepción. El uso de mutantes ha sido de gran utilidad para determinar su función; sin embargo, aún falta mucho por estudiar. Los resultados preliminares del efecto de los brasinoesteroides en la inducción de la división celular probablemente nos lleven a nuevas áreas del conocimiento. Se está conociendo cada vez más cuáles son los componentes intermediarios para llevar la señal de la membrana a la célula, pero aún quedan muchas preguntas por responder. Por otro lado, aunque se ha comprobado que BRI1 es el receptor de los brasinoesteroides, se han reportado otras mutantes insensibles, lo que puede llevar al descubrimiento de otro tipo de receptores. Además, BRI1 también podría ser el receptor de una sustancia de origen proteico denominado sistemina, con propiedades similares a las hormonas vegetales. Dados los esfuerzos que se están realizando por parte de diferentes grupos de trabajo en todo el mundo, se espera que en un tiempo razonablemente corto se tengan definidos claramente sus mecanismos y modo de acción. ![]()
Bibliografía
Adam, G. y V. Marquardt (1986), “Brassinosteroids”, Phytochemistry, 25 (8), 1787-1799.
Azpeitia, A., J. L. Chan, L. Sáenz y C. Oropeza (2003), “Effect of 22(S), 23(S)-homobrassinolide on somatic embryogenesis in plumule explants of Cocos nucifera (L.) cultured in vitro”, Journal of Horticultural Science and Biotechnology, 78, 591-596.
Bishop, G. J., y T. Yokota (2001), “Plant steroid hormones, brassinosteroids: current highlights of molecular aspects on their synthesis/metabolism; transport, perception and response”, Plant Cell Physiology, 42(2), 114-120.
Buchanan, B. B., W. Gruissen, R. L. Jones (comps.) (2000), Biochemistry and molecular biology of plants, Maryland, eua, American Society of Plant Physiologist.
Mandava, n. b. (1988), “Plant growth-promoting brassinosteroids”, Annual Review of Plant Physiology and Plant Molecular Biology 39, 23-52.
Nemhauser, J.L., y J. Chory (2004), “Bring it on: new insights into the mechanism of brassinosteroid action”, Journal of Experimental Botany 55 (395), 265-270.
Luis Sáenz es licenciado en química por la Facultad de Química de la Universidad Autónoma de Yucatán. Estudió la maestría y el doctorado en el Centro de Investigación Científica de Yucatán. Posteriormente realizó una estancia posdoctoral en el Institut de Recherche pour le Développement en Montpellier, Francia. Ha publicado varios artículos científicos en revistas internacionales y nacionales. Actualmente es investigador del Centro de Investigación Científica de Yucatán. Su investigación está enfocada a estudiar la regulación del ciclo celular durante el proceso de embriogénesis somática en plantas.
Esta dirección de correo electrónico está siendo protegida contra los robots de spam. Necesita tener JavaScript habilitado para poder verlo.
Iván Córdova obtuvo su licenciatura en biología en la Universidad Autónoma de Yucatán. Estudió la maestría en Biotecnología de Plantas en el Centro de Investigación Científica de Yucatán. Posteriormente llevó a cabo una estancia de un año en el Instituto de Rothamsted, en Inglaterra. Ha publicado varios artículos científicos en revistas nacionales e internacionales. Actualmente trabaja en el Centro de Investigación Científica de Yucatán como técnico académico.
Esta dirección de correo electrónico está siendo protegida contra los robots de spam. Necesita tener JavaScript habilitado para poder verlo.
Francisco Rodríguez obtuvo el título de ingeniero agrónomo especialista en fitotecnia por la Universidad Autónoma de Chapingo. Actualmente realiza estudios de maestría en el Centro de Investigación Científica de Yucatán, donde aborda el efecto de los brasinoesteroides sobre la expresión de genes que regulan el ciclo celular durante la embriogénesis somática en cocotero. Es becario del Consejo Nacional de Ciencia y Tecnología.
Esta dirección de correo electrónico está siendo protegida contra los robots de spam. Necesita tener JavaScript habilitado para poder verlo.