| El c o n t r o l |
| del ciclo celular |
| en plantas |
| |
Juan G. Colli-Mull,
Luis Carlos Rodríguez Zapata, Enrique Castaño de la Serna |
|


|
|
Introducción
El crecimiento y desarrollo de los organismos dependen de la coordinación entre el crecimiento y la multiplicación de sus células. En los organismos unicelulares, la división celular implica una verdadera reproducción, ya que por este proceso se generan dos células. Por el contrario, los organismos multicelulares derivan de la fusión de dos células que forman un cigoto, y la repetida división de esta célula y sus descendientes determinan el crecimiento y desarrollo del individuo.
El ciclo celular es un conjunto ordenado de eventos que culmina con la división de una célula madre en dos células hijas idénticas. La división celular es una de las características más notables de la vida, y los mecanismos generales que la controlan son comunes a todos los organismos eucariontes, incluyendo protozoarios, hongos, plantas y animales. Esto se debe a que los organismos eucariontes tienen en común un gran número de proteínas, las cuales se encuentran conservadas evolutivamente (es decir, son similares entre distintas especies) tanto en función como, en muchos de los casos, en secuencia (el orden en que están unidas las unidades que las forman, los aminoácidos). Tal es el caso de varias proteínas requeridas para controlar el ciclo celular.
El ciclo celular de los eucariontes está regulado en múltiples puntos, en los que se requiere la activación de una familia de enzimas llamadas proteínas cinasas de serina/treonina. Éstas necesitan para su actividad de la unión de una proteína regulatoria conocida como ciclina, de la cual surge su nombre de cinasas dependientes de ciclinas o CDK.
En los eucariontes, el ciclo celular está definido por cuatro fases, denominadas: G1, S (en la que se realiza la síntesis del ácido desoxirribonucleico, o ADN), G2 y M (durante la cual ocurre la división celular, o mitosis).
La fase G1, que precede a la fase S, se caracteriza por ser el principal punto de regulación por estímulos extracelulares dados por reguladores del crecimiento. Durante esta fase la célula queda habilitada para producir todas las proteínas necesarias para la replicación del ADN. Durante la etapa temprana de la fase G1 se revisa el estado metabólico de la célula antes de avanzar hasta una etapa de compromiso en el ciclo, definida por un punto en el que la célula "decide" si se compromete o no a completar el ciclo celular.
Unas vez que pasa este punto, la replicación del ADN se realiza en la denominada fase S; ésta es seguida por otra revisión conocida como fase G2, donde se ejercen controles moleculares que aseguran que las dos nuevas cadenas de ADN se encuentren intactas antes de entrar a la fase de separación cromosómica, o fase M, en la cual se lleva a cabo la citocinesis, fenómeno por medio del cual se obtienen dos células hijas. Una vez completado el ciclo, la célula puede comenzar nuevamente otra ronda de división, diferenciarse, morir, o salir del ciclo celular, según las condiciones existentes.
Factor transcripcional E2F
La molécula conocida como E2F, factor involucrado en la activación del promotor viral E2, representa una familia de proteínas, la cual en humanos comprende siete miembros (E2F1 a E2F7) los cuales encuentran pareja con alguno de los dos miembros de la proteína DP (Trimarchi y Lees, 2002). E2F fue caracterizado originalmente como una actividad celular requerida para la expresión de la proteína E1A del adenovirus, que interviene en la activación transcripcional del promotor viral E2. Estudios posteriores demostraron que E2F, al unirse al ADN, controla la transcripción de genes esenciales para la división celular. Se han identificado proteínas tipo E2F y DP en varias especies de plantas, incluyendo trigo, tabaco, zanahoria, Arabidopsis y arroz (Wen-Hui Shen, 2002).
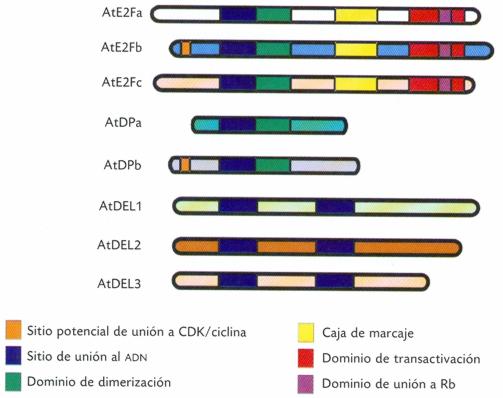
Con la publicación del genoma de Arabidopsis (Arabidopsis Genome Initiative, 2000) se ha identificado la presencia de seis E2F y dos DP. Lo interesante es que mientras atE2Fa-c necesitan formar heterodímeros con atDPs para lograr una unión eficaz al ADN y regular la expresión génica, los otros tres miembros de la familia de E2F, los cuales tienen duplicado el dominio (región de la molécula de proteína) de unión al ADN, se unen eficientemente en ausencia de atDPs. Además, las proteínas E2F que han sido ya caracterizadas en Arabidopsis y otras especies de plantas contienen dominios similares a las proteínas de E2F en animales, incluyendo un dominio de dimerización, un dominio de transactivación, un posible sitio de unión de CDKs en E2Fb y DPb y varios sitios probables de fosforilación en la familia de E2F, los cuales están involucrados en la regulación de su actividad transcripcional (Figura 1).
La ruta E2F/PRB en el control de la transición de G1a S
Se ha propuesto un modelo del control de la transición G1-S en el ciclo celular en el cual las ciclinas tipo D (CycD) son los mediadores primarios de la transición y tienen la responsabilidad de estimular el avance del ciclo celular. La transcripción de las CycD es activada por señales extracelulares, lo cual lleva a la formación de complejos activos CDK-CycD. Este complejo activo CDK-CycD es capaz de fosforilar e inactivar a la proteína del retinoblastoma (pRb) un factor antiproliferativo, la cual se separa de E2F, permitiendo así la activación de genes regulados por E2F. Cabe mencionar que la fase G1 del ciclo celular generalmente representa el mayor punto de regulación de las fases restantes.
|
|
|
Figura 1. Organización estructural y propiedades de la familia de E2F en Arabidopsis. En la familia de E2F se conservan los dominios de unión al ADN, de dimerización, de caja de marcaje y el dominio de transactivación. Basado en la conservación de los diferentes dominios, las ocho proteínas de E2F de Arabidopsis son clasificadas en E2F, DP y DEL.
|
|
Una ruta análoga de E2F/pRb en la transición G1-S del ciclo celular, que involucra la fosforilación de pRb por los complejos CDK/ciclina y la liberación de E2F, ha sido dilucidada recientemente en células vegetales, y probablemente los mecanismos de su regulación sean los mismos que en sus contrapartes en humanos y otros eucariontes superiores (Figura 2). Aunque no se han encontrado homólogos de las ciclinas E en plantas, fundamentales en la transición G1-S en animales, las ciclinas tipo A o quizá algunas del tipo D podrían participar en la regulación de la transición G1-S y de G2-M del ciclo celular (Wen-Hui Sen, 2001).
Durante la fase G1, la proteína Rb se encuentra en su forma activa (hipofosforilada) y puede unirse a activadores transcripcionales de la familia de E2F, dando como resultado la formación de un complejo que reprime la transcripción de genes clave para E2F y, consecuentemente, impide la progresión de la fase G1/S. Conforme avanza la fase G1, pRb es fosforilada secuencialmente por diferentes complejos CDK/ciclinas, inactivándola y finalmente liberando a E2F para activar genes requeridos para el avance del ciclo celular (Figura 2).
|
|
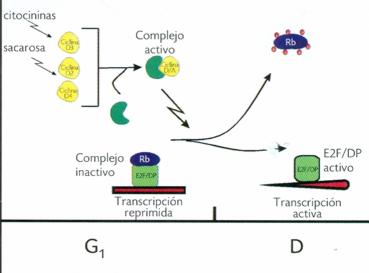
Figura 2. Modelo propuesto para la regulación de la transición G1-S del ciclo celular en plantas. La expresión de las ciclinas D es estimulada por señales del medio ambiente y de crecimiento. Estas ciclinas se pueden asociar a la CDK-A. Este complejo activo ciclina D/CDK-A puede fosforilar a la Rb, la cual se encuentra formando un complejo con E2F/DP, inhibiendo la transcripción. Conforme avanza el ciclo celular, Rb es hiperfosforilada por el complejo ciclina D/CDK-A en G1 tardía, lo que ocasiona que se libere el complejo transcripcional E2F/DP, el cual es capaz de activar la transcripción de genes requeridos para la fase S del ciclo celular.
|
|
Genes regulados por E2f que participan en el ciclo celular
Los niveles de transcripción de E2F en plantas son generalmente bajos. Sin embargo, se ha demostrado en Arabidopsis que hay una expresión diferenciada, dependiendo del tejido. Por otra parte, en suspensiones celulares parcialmente sincronizadas los niveles de transcripción de E2F muestran un incremento en la transición de G1 a S del ciclo celular (Ramírez-Parra y colaboradores, 1999).
El control en la transición G1-S generalmente representa el punto crucial en la división celular. En la fase G1 del ciclo celular las células "deciden" si permanecen o salen del ciclo mitótico. A este punto de control se le ha llamado Start, en levaduras, y "punto de restricción" (R), en animales. Aunque tales puntos no han sido definidos exactamente en plantas, la aplicación de inhibidores químicos reveló un punto crucial de control en la fase G1 en células de tabaco (Wen-Hiu Shen, 2001).
Así, E2F juega un papel primordial en el control de genes requeridos para el avance del ciclo celular. Entre estos genes se encuentran el de la ciclina A, genes de histonas y genes involucrados en la síntesis de ADN, como la ADN polimerasa ?, la dihidrofolato reductasa, CDC6, ORC1, ORC3, RPA, MCM, el antígeno nuclear de proliferación celular (PCNA), la ribonucleótido reductasa y una subunidad de la polimerasa ? (Ramírez-Parra y colaboradores, 2003).
|
|
E2F regula una variedad de genes funcionales, no sólo los relacionados al ciclo celular
Recientemente se ha demostrado que el complejo E2F/DP juega un papel importante para regular la expresión de genes, no sólo en células en estado proliferativo, sino también durante la diferenciación, el desarrollo y en procesos de apoptosis (muerte celular programada; Muller y colaboradores, 2001; Braceen y colaboradores, 2004).
En la última década, debido al gran avance de la investigación sobre la regulación del ciclo celular en las plantas, es posible comprender más a fondo la regulación de este proceso vital, en el que los componentes principales son similares a los presentes en los animales. Sin embargo, las plantas poseen características únicas de crecimiento y desarrollo, lo que trae por consiguiente que hayan desarrollado estrategias particulares en el control del ciclo celular, generando nuevas rutas de desarrollo.
Estudios realizados en Arabidopsis demostraron que en los ápices foliares de esta planta se encuentran varios tipos de tejidos que difieren tanto en su estado de crecimiento como de diferenciación. En tejidos que se dividen activamente, en los primordios foliares, en tejido vascular de los primordios maduros, en botones axilares y en la epidermis y dermis del hipocotilo se encuentra un alto nivel de expresión de E2Fa/DPa, lo que sugiere que E2Fa-DPa no sólo regula el avance del ciclo celular mitótico, sino que también participa en la diferenciación celular (Lieven de Veylder y colaboradores, 2002). Recientemente Ramírez-Parra y colaboradores (2003) realizaron una investigación manejando el genoma de Arabidopsis para encontrar genes que pudieran ser regulados por E2F/DP. En este estudio se enfocaron en un sitio bien definido de unión de E2F al ADN, así como en un sitio variante del mismo. Este sitio de unión al ADN se encuentra altamente conservado entre humanos y plantas, incluyendo Arabidopsis, en particular la hélice ?3 involucrada directamente en el contacto con el ADN (Zheng y colaboradores, 1999).
Los análisis dieron como resultado la identificación de 126 genes que contienen el motivo de unión E2F al ADN en la región promotora de estos genes, los que fueron clasificados en las siguientes categorías funcionales: genes que participan en el ciclo celular y en la reparación del ADN (31.3 por ciento) y genes que participan en la transcripción (21.7 por ciento) -en estas categorías se encuentra el mayor porcentaje de genes expresados-; además se encontraron, con menores porcentajes, genes involucrados en los procesos de estrés y defensa, en transducción de señales, en organización del citoplasma, en transporte, en metabolismo y en biogénesis celular (Ramírez-Parra et al., 2003)
Ahora bien, no olvidemos que los resultados anteriores son solamente estudios preliminares, que utilizan aproximaciones geonómicas para identificar genes regulados por E2F. Por lo anterior, los estudios futuros podrían enfocarse a genes específicos para demostrar la función de E2F en la regulación de su expresión.
A pesar de las grandes interrogantes que aún persisten sobre la regulación de la división celular en plantas, la evidencia actual permite concluir que éstas utilizan, en las vías que regulan la división celular, mecanismos similares a los que se encuentran en animales. Es de esperarse, sin embargo, que el rápido avance de la investigación en este campo de la ciencia arroje, en un futuro próximo, una visión más clara de la regulación del ciclo celular vegetal.
|
|
|
| |
|
Juan Gualberto Colli-Mull es licenciado en Biología por el Instituto Tecnológico Agropecuario No. 2, y maestro en Ciencias y Biotecnología de plantas por el Centro de Investigación Científica de Yucatán (cicy). Actualmente está cursando el doctorado en Ciencias y Biotecnología de Plantas en el CICY. Su trabajo se centra en el estudio de la función de E2F en procesos de desarrollo y apoptosis en cultivos in vitro de Coffea canephora. Esta dirección de correo electrónico está siendo protegida contra los robots de spam. Necesita tener JavaScript habilitado para poder verlo.
Luis Carlos Rodríguez Zapata es investigador del cicy. Cursó la licenciatura en Biología de la Universidad Autónoma de Yucatán y el doctorado en Ciencias y Biotecnología de plantas en el CICY. Realizó entrenamientos posdoctorales en el Instituto de Biotecnología de la Universidad Nacional Autónoma de México (unam) y en la Universidad Estatal de Nueva York, en Stony Brook. Su interés se centra en la transformación de cultivares de plátano. Esta dirección de correo electrónico está siendo protegida contra los robots de spam. Necesita tener JavaScript habilitado para poder verlo.
Enrique Castaño de la Serna es maestro y doctor en Ciencias por la Universidad de Rochester, Nueva York. Realizó entrenamientos posdoctorales en la Universidad de Harvard y en el Instituto Marie Curie, en Inglaterra. Actualmente es investigador del CICY. Su línea de investigación radica en los estudios mecanísticos del control transcripcional. enriquec@cicy.mx
|
| |
|
Bibliografía
Bracken, Adrian P., Marco Ciro, Andrea Cocito y Kristian Helin (2004), "E2F target genes: unraveling the biology", Trends in Biochemical Science, 29, 409-417.
De Veylder, Lieven, T. Beeckman, G. T. S. Beemster, J. de Almeida, E. S. Ormenese, S. Maes, M. Naudts, E. Van Der Schueren, A. Jacqmard, G. Engler y Dirk Inze (2002), "Control of proliferation and of reduplication and differentiation by the Arabidopsis E2Fa-DPa transcription factor", The EMBO Journal, 21, 1360-1368.
Müller, H., A. P. Bracken, R. Vernell, M. C. Moroni, F. Christians, F. Grassilli, E. Prosperini, E. Vigo, J. D. Oliner y K. Helin (2001), "E2Fs regulate the expression of genes involved in differentiation, development, proliferation and apoptosis", Genes Dev., 15, 267-285.
Ramírez-Parra, E., Xie, Q, Boniotti, M. B. y C. Gutiérrez (1999), "The cloning of plant E2F, a retinoblastoma-binding protein, reveals unique and conserved features with animal G1/S regulators", Nucleic Acids Research, 27, 3527-3533.
Ramírez-Parra, E, Corinne Frundt y C. Gutiérrez (2003), "A genome-wide identification of E2F-regulated genes in Arabidopsis", The Plant Journal, 33, 801-811.
Trimarchi, J. M. y J. A. Lees (2002), "Sibling rivaly in the E2f family", Nat. Rev. Mol Cell Biol., 3, 11-20.
Wen-Hui Shen (2002), "The plant E2F-Rb pathway and epigenetic control", Trends in Plant Science, 7, 505-511.
Wen-Hui Shen (2001), "The plant cell cycle: G1/S regulation", Euphytica, 118, 223-232.
Zheng N., N, E. Fraenkel, C. O. Pabo y N. P. Pavletich (1999), "Structural basis of DNA recognition by the heterodimeric cell cycle transcription factor E2F-DP, Genes Dev., 13, 666-674.
|
| |
|
|
|




