El desierto y sus habitantes: el caso de la hormiga roja
Introducción
La zona semiárida conocida como el valle de Zapotitlán de las Salinas, en el estado de Puebla, es un lugar mágico; un claro ejemplo de cómo la fuerza de la tectónica de placas ejerce su influencia sobre la fisonomía de la faz del planeta, si consideramos que hace más de 65 millones de años, cuando los dinosaurios aún poblaban la Tierra, este lugar era el fondo de un lecho marine Pétreos testigos, como fósiles de caracoles diversos y de corales, dan fe de un húmedo pasado.
Esta región es única en términos de riqueza biológica, pues cerca del 30 por ciento de sus especies son endémicas; es especialmente rica en cactáceas columnares. Más aún, la vegetación que actualmente cubre el valle posiblemente se ha mantenido sin muchos cambios desde hace 10 mil años (Smith, 1967). Sin embargo, el valle de Zapotitlán, como el resto de las zonas áridas, es un ambiente estresante, limitado por la esporádica e impredecible precipitación (de 300 a 400 milímetros de lluvia anual) y agobiado por la temperatura que puede rebasar los 70 grados centígrados al nivel del suelo. Estos factores ocasionan que la productividad primaria esté tres órdenes de magnitud por debajo de la de otros ecosistemas, favoreciendo la idea equivocada de que los desiertos son sistemas simples marcados por su escasa biodiversidad, tanto en número de especies como en cantidad de interrelaciones bióticas (entre organismos).
Las hormigas, grupo diverso y sensible a los cambios ambientales, son un componente importante en esta isla de variabilidad biológica y condiciones hídricas estresantes, tanto por su abundancia como por los papeles ecológicos que desempeñan. Entre las especies que más destacan en Zapotitlán por su abundancia y fácil observación en el campo, se encuentran Atta mexicana (hormiga arriera), Camponotus rubrithorax (ixcamel) y Pogonomyrmex barbatus (hormiga roja), aunque hay más de diez especies registradas en el sitio (Ríos-Casanova y colaboradores, 2004; Figura 1).
¿Qué es la productividad primaria?
La productividad primaria es la cantidad de tejido vegetal que pueden producir las plantas y que mantiene a los consumidores primarios (herbívoros); éstos, a su vez, mantienen a los secundarios (carnívoros). A pesar de que existe un debate aún no resuelto acerca de la forma de medirla, la productividad primaria de un ecosistema ha sido entendida en términos de biodiversidad vegetal y precipitación. Dicha relación se hace más evidente cuando se manejan escalas geográficas amplias, que permiten comparar ecosistemas completos. Un ecosistema productivamente rico podría estar representado por una selva alta perennifolia (que nunca pierde su follaje, como la selva lacandona); uno relativamente pobre, por un desierto (como el chihuahuense).
Aunque este razonamiento tiene una fuerte base empírica, ya que la idea es que el agua limita la productividad primaria y ésta, a su vez, la energía disponible para los consumidores, controlando la productividadsecundaria, la evidencia indica que los desiertos son sitios sorprendentemente diversos respecto a ciertos grupos de plantas y animales, con una fuerte heterogeneidad espacio-temporal, y una variedad de interacciones bióticas, que son a su vez, factores que promueven la complejidad (Polis, 1991). De esta manera, una definición que tenga como base la diversidad y precipitación como indicadores de productividad no soluciona el problema de explicar la biodiversidad de los desiertos. El principal problema es que al considerar a las zonas áridas como lugares pobres, estamos diciendo que son en consecuencia sitios poco diversos y simples. Esto se contrapone a lo evidente y no explica la enorme riqueza biológica de la que México es depositario, con el 60 por ciento de su territorio ocupado por zonas xéricas (con deficiencia de agua).

Figura 1. Especies comunes de hormigas en el valle de Zapotitlán de las Salinas. Fotografía: Laboratorio de Microartrópodos Terrestres, Facultad de Ciencias, UNAM.
Además, cabría un pensamiento opuesto al fundamento empírico de la idea de que la humedad limita la diversidad y la productividad. Los desiertos, al ser ambientes inhóspitos dominados por factores abióticos (temperatura y humedad, principalmente), podrían propiciar que los organismos respondan de formas distintas a las condiciones existentes, favoreciendo el origen de especies nuevas y la biodiversidad, como evidente-mente ha ocurrido en Zapotitlán y en otros desiertos de México (Figura 2; Valiente-Banuet, 1990; Briones, 1994). Así, la limitación de los recursos, promovida por una baja productividad, puede conducir al surgimiento de estrategias adaptativas que originen una alta riqueza de especies (Schowalter, 2000).
Algunas investigaciones han concluido que la diversidad no está relacionada linealmente con la productividad, porque sitios con niveles intermedios de productividad a menudo soportan una alta diversidad, y una productividad mayor favorece la dominancia de especies más competitivas. Lo anterior se ajusta a la hipótesis del disturbio intermedio, que establece una relación no lineal entre productividad y diversidad (Wilkinson, 1999).
¿Cómo se adaptan los organismos a las condiciones áridas?
Las adaptaciones de los organismos que viven en las zonas áridas tienen que ver con el aprovechamiento del agua y la obtención de alimento. Las plantas han desarrollado la presencia de cutículas gruesas, densas capas de pubescencia, colores verdes blanquecinos que reflejan la luz, hojas de tipo escamoso y la presencia de tejidos suculentos que, además de constituir reservorios de agua, favorecen el desarrollo de la vía fotosintética CAM (Metabolismo Ácido de las Crasuláceas) (Valiente-Banuet, 1990).
Asimismo, las cactáceas, características de estos ambientes, ajustan la orientación e inclinación de sus estructuras en función del efecto que ejerce la luz sobre ellas, con la finalidad de captar la radiación fotosintética necesaria para realizar la fijación de dióxido de carbono, el aumento de peso seco y una floración exitosa, sin poner en riesgo su estado físico. Las plantas anuales han adaptado su forma de vida de modo que sólo germinan y florecen durante cortos periodos de tiempo, cuando la humedad es adecuada para su crecimiento. Este es un caso interesante, ya que las lluvias en los desiertos no son uniformes en el tiempo ni en el espacio. La precipitación que cae en un lugar no es homogénea para toda el área, y este fenómeno genera que las comunidades de plantas anuales fluctúen en abundancia y producción de semillas, haciendo que las especies no se presenten continuamente año con año. Esto contribuye a la formación de manchones de distribución que cambian constantemente.
Por su parte, animales como los mamíferos están activos durante la noche o en las horas de menos calor, y tienen pelajes claros. Los insectos, como las hormigas, modifican sus estrategias de forrajeo en función de la disponibilidad y variación espacial en la que se presentan los recursos (Gordon, 1991), construyen graneros dentro de sus nidos (como P. barbatus), cultivan hongos utilizando hojas y otras estructuras vegetales como sustrato (como A. mexicana) o establecen relaciones mutualistas con algunas plantas (C. rubrithorax y Ferocactus latispinus variedad spiralis). Así confirman la capacidad que han tenido para ocupar una gran variedad de nichos, su habilidad de adaptarse rápidamente a los cambios del ambiente (Schowalter, 2000) y su influencia sobre la abundancia y composición de las comunidades vegetales. Esto, además del papel ecológico que tienen en los ecosistemas, hace que las hormigas sean organismos importantes desde el punto de vista biológico: las asociaciones que mantienen con cerca de 465 especies de plantas de 52 familias, y con artrópodos (áfidos, orugas, arañas, colémbolos, escarabajos, moscas, saltamontes, otras hormigas, etcétera). En su mayoría se trata de relaciones de tipo mutualista (mutua ventaja). También son dispersoras y consumidoras importantes de semillas, además de comer vegetales (herbívoras), hongos (fungívoras) y ser incluso depredadoras de otras hormigas (Schultz y McGlynn, 2000).
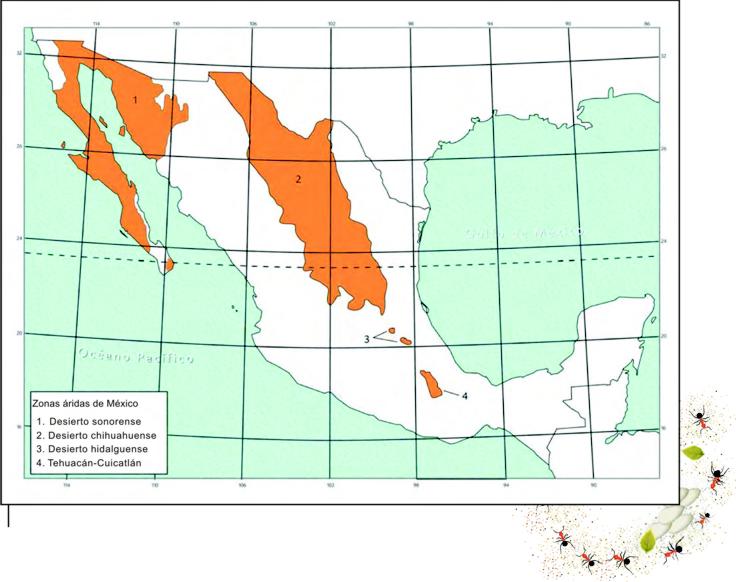
Figura 2. Distribución de zonas áridas en México (basado en Briones, 1994).

Actividad de hormigas rojas (Pogonomyrmex barbatus) sobre el nido. Fotografía: Rafael Guzmán.
La hormiga roja granívora en Zapotitlán de las Salinas, Puebla
La hormiga roja (Pogonomyrmex barbatus) pertenece a un género cuyas especies se distribuyen por todo el continente americano. Cerca de la mitad son cosechadoras o granívoras -términos empleados para los organismos que consumen semillas. Se encuentran principalmente en hábitats áridos, pueden tener una longevidad de 15 a 50 años (McMahon y colaboradores, 2000) y son fáciles de encontrar en Zapotitlán. Se ha calculado que en una hectárea pueden encontrarse hasta 180 mil hormigas rojas buscando alimento y, como las semillas son la base de su dieta, esto las convierte en actores primordiales en la dinámica de la comunidad de plantas, pues modifican la abundancia y composición de sus especies.
Sin embargo, debido a que las plantas anuales son un componente importante de la comunidad y a que éstas dependen de las impredecibles e inconstantes lluvias, pueden a su vez determinar la estructura de las poblaciones y comunidades no sólo de las hormigas granívoras, sino también de otros grupos de organismos, como aves y roedores, que dependen de las semillas. Las marcadas fluctuaciones espaciotemporales en la disponibilidad de los recursos que se presenta en los desiertos hace interesante observar cómo responden los granívoros ante esta situación de incertidumbre.
 |
 |
|
Microambientes dentro del Jardín Botánico Helia Bravo, en Zapotitlán de las Salinas. a) Llano: el sitio menos diverso. b) Jardín: el sitio con mayor diversidad vegetal. Fotografías: J. Alejandro Zavala y Rafael Guzmán. |
Cuando se compara el patrón de forrajeo en dos microambientes diametralmente diferentes en cuanto a diversidad vegetal (ver fotografías), y esto a su vez se toma como indicador de la abundancia y variedad de alimento disponible para las hormigas, se encuentran patrones realmente interesantes. La fenología (la medida en que las plantas producen tejido vegetal, como hojas, flores o frutos) y la composición de la comunidad vegetal juegan un papel importante. Los análisis comparativos de los hábitos alimentarios y de actividad de la hormiga roja señalan que, en un sitio más diverso, las hormigas no necesariamente son más activas como resultado de la mayor disponibilidad de recursos. De ello podría suponerse el efecto de la producción de más cantidad y variedad de semillas durante los meses de lluvia. Además, el espectro trófico (las preferencias alimentarias) es más amplio durante esta época del año en un sitio diverso que en uno depauperado y menos diverso (Figura 3).
El hecho de que, a pesar de la enorme diversidad que pueda poseer un lugar, no haya semillas disponibles, es prueba de que la fenología de las plantas que constituyen la comunidad vegetal de ese lugar juega un papel importante en la dieta de las hormigas (como lo pueden jugar también otros factores: competencia y depredación), pues la modifican considerable mente y provocan que se vuelvan más generalistas en su alimentación.
Una observación detallada del estado fenológico de las plantas del valle de Zapotitlán indicó que después de la época de lluvias, ya cuando las plantas anuales se han secado, las semillas disponibles son principalmente las de plantas perennes, como biznagas (Mamillaria carnea, Ferocactus lastispinus variedad spiralis, Ferocactus robustus), agaves (Agave marmorata, A. macroacantha, A. karwinskii), lechuguillas (Hechita podantha), cactos (Opuntia pillifera, O. decumbens, Myrtillocactus geometrizans, Equinocactus platyachantus), asteráceas (Sanvitalia fruticosa) y árboles (Ipomoea arborecens, Fouquieria formosa). Esto modifica la amplitud del espectro alimentario y la actividad de los granívoros que viven en sitios donde estas plantas son el componente dominante de la vegetación durante la época seca del año (Guzmán, datos no publicados).
|
|
|
Figura 3. Comparación de la amplitud de las preferencias en la utilización de recursos por las hormigas durante la estación lluviosa en Zapotitlán de las Salinas. Un valor del índice de selección cercano a uno indica una alta preferencia por el recurso. |
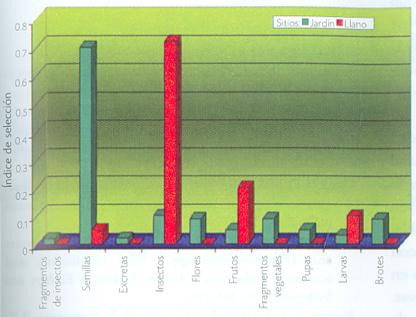
Estos resultados fundamentan la discusión anterior de que la diversidad en sí misma, tomada como el número de especies diferentes, no es indicador lineal de productividad, sino qué especies existen y cuál es la abundancia de cada una de ellas dentro de la comunidad vegetal. Esta conclusión coincide con Naeem y colaboradores (1999), quienes comprueban la importancia de conocer la composición y la identidad de las especies que integran la comunidad de plantas, debido a que pueden variar dramáticamente en su contribución al funcionamiento del ecosistema.
Por otro lado, la flexibilidad de conductas en la búsqueda de alimento es de vital importancia cuando la disponibilidad de los recursos no es constante. Las hormigas rojas, así como las Ectatommini (por ejemplo, las Ectatomma ruidum; Franz y Weislo, 2003), deben de tener además una serie de mecanismos de aprendizaje y memoria que les permitan forrajear eficientemente en los lugares donde han encontrado alimento.
Uno de estos mecanismos podría estar relacionado con las grandes distancias que son capaces de recorrer (más de 30 metros), sobre todo en los sitios donde la vegetación perenne es escasa y la vegetación anual predomina durante los cortos periodos de lluvia. Las hormigas, al ser altamente exploradoras, deben de tener un mapa mucho más complejo de la distribución de las colonias vecinas, y aun de las que están más alejadas de sus proximidades, de lo que en un principio un observador podría pensar. Pero, más allá de esto, es muy probable que además de tener una idea compleja de la distribución de sus vecinas cercanas y distantes, también tengan información acerca de los lugares donde sea posible encontrar alimento mucho más allá de sus fronteras. Lo antedicho proporciona evidencia de que bajo ciertas circunstancias (sobre todo de escasez), las fronteras de las colonias no están fuertemente limitadas. Por el contrario, la forma de los territorios sería semejante a la de los seudópodos de las amibas, causando que las forrajeras de una u otra colonia se encuentren continuamente sin que esto implique el inicio de hostilidades entre ellas. A su vez, lo anterior promovería la coexistencia en un ambiente difícil e impredecible (Gordon, 1991).
En conclusión
Las condiciones ambientales que gobiernan en los desiertos (temperatura, precipitación y suelo) propician que estos ecosistemas no sean homogéneos; es decir, que el paisaje esté constituido de parches con diferente composición vegetal, que varían en el espacio y el tiempo. Esta variabilidad, en el marco de la austeridad ambiental de las tierras áridas, tiene importancia debido a que ligeras irregularidades en un componente del ecosistema, como la precipitación, propician variaciones grandes en otros componentes, creando distintos microambientes (Wilby y Shachak, 2000). Por ejemplo, diferencias en la erosión, patrones de drenaje, composición del sustrato, viento, e influencias biológicas (como la actividad de construcción de nidos por parte de las hormigas) pueden ocasionar que la distribución de los nutrientes sea extremadamente irregular, en parches (Polis, 1991), lo cual afecta drásticamente la distribución y abundancia de plantas, la productividad, la composición y abundancia de los consumidores primarios y sus depredadores (Polis, 1991).
Por otro lado, se favorece que los organismos respondan de formas diversas a las condiciones existentes, impulsando el origen de especies nuevas, la biodiversidad y un amplio espectro de interrelaciones entre especies (Polis, 1991), como posiblemente ha ocurrido en Zapotitlán y en otros desiertos de México, de manera que la limitación de los recursos promovida por una baja productividad puede conducir al surgimiento de estrategias adaptativas que originen una alta riqueza de especies (Schowalter, 2000).
Otro aspecto interesante es que la productividad no puede ser medida en términos de la diversidad vegetal, sino de la identidad y la composición de la comunidad, ya que no todas las plantas contribuyen por igual al funcionamiento del ecosistema. Es posible, entonces, que se tengan lugares muy diversos pero con escasa productividad, y lugares poco diversos pero en los que una o dos especies sean altamente productivas. Por ello, se debe ser muy cuidadoso al momento de establecer una relación tan lineal como ésta.
Agradecimientos
Quiero agradecer a Águeda Vásquez Sosa, María del Carmen Herrera Fuentes y José Alejandro Zavala Hurtado por sus comentarios y ayuda en la redacción del texto; al revisor anónimo, quien proporcionó sugerencias importantes para enriquecer este trabajo; a Pedro Luis Guzmán Álvarez, por su apoyo, y a Gabriela Castaño Meneses, por facilitar las fotografías de la Figura 1.
Bibliografía
Briones, V. O. L. (1994), "Origen de los desiertos mexica-nos", Ciencia, 45, 263-279.
Franz, N. M. y W. T., Wceislo (2003), "Foraging behaviour in two species of Ectatomma (Formicidae: Ponerinae): individual learning of orientation and timing", Journal of Insect Behaviour, 16, 381-410.
Gordon, D. M. (1991), "Behavioral flexibility and the foraging ecology of seed-eating ants", The American Naturalist, 138, 379-411.
McMahon, J. A., J. F. Mull y T. O. Crist (2000), "Harvester ants (Pogonomyrmex spp.): their community and ecosystem influences", Annual Review of Ecology and Systematics, 31, 265-291.
Naeem, S., F. S. Chair, F. S. Chapin III, R. Constanza, P. R. Ehrlich, F. B. Golley, D. U. Hooper, J. H. Lawton, R. V. O'Neill, H. A. Mooney, O. E. Sala, A. J. Symstad y D. Tilman (1999), "Biodiversity and ecosystem functioning: maintaining natural life support processes", Issues in Ecology, 4, 2-11.
Polis, G. A. (1991), "Desert communities: an overview of patterns and process", en G. A. Polis (comp.), The Ecology Desert Communities, Tucson, The University of Arizona Press, pp. 1-26.
Ríos-Casanova, L., A. Valiente-Banuet y V. Rico-Gray (2004), "Las hormigas del valle de Tehuacán: comparación con otras zonas áridas de México", Acta Zoológica Mexicana, 20, 37-54.
Schowalter, T. D. (2000), Insect Ecology: An Ecosystem Approach, San Diego, California, Academic Press.
Schultz, T. R. y T. P. McGlynn (2000), "The interactions of ants with other organisms", en Agosti D., J. D. Majer, L. E. Alonso y T. R. Schultz (comps.), Ants: Standard Methods for Measuring and Monitoring Biodiversity, Washington, Smithsonian Institution Press, pp. 35-44.
Smith Jr., C. E. (1967), "Plant remains", en D. S. Byers (comp.), The Prehistory of the Tehuacan Valley. Environment and subsistence, vol. 1, Austin, Texas, Universidad de Texas, pp. 220-225.
Valiente-Banuet, A. (1990), "Los desiertos de México", Revista de la Sociedad Mexicana de Historia Natural, 41, 83-84.
Wilby A. y M. Shachak (2000), "Harvester ant response to spatial and temporal heterogeneity in seed availability: pattern in the process of granivory", Oecologia, 125(4), 495-503.
Wilkinson D. M. (1999), "The disturbing history of intermediate disturbance", Oikos, 84, 145-147.
Rafael Guzmán Mendoza es maestro en ciencias con especialidad en ecología de insectos sociales de la familia Formicidae y profesor-investigador de la Universidad Intercultural del Estado de México, en la División de Desarrollo Sustentable. Ha publicado varios artículos especializados y de divulgación dentro del área de ecología de insectos sociales, y recibió por parte de la Universidad Autónoma Metropolitana, el Premio al Mérito Universitario por su tesis de maestría. Esta dirección de correo electrónico está siendo protegida contra los robots de spam. Necesita tener JavaScript habilitado para poder verlo.