Las selvas tropicales y el cambio climático
![]()
Introducción
El incremento en la concentración atmosférica de dióxido de carbono (CO2) generado a raíz de la revolución industrial, ha ocasionado cambios en la concentración atmosférica de este gas, que ha aumentado de 280 partes por millón (una parte por millón equivale a un miligramo de sustancia por litro en agua, o un gramo en mil litros), durante el periodo preindustrial, a 370 partes por millón en la actualidad. Si las actividades humanas continúan incrementando las emisiones de dióxido de carbono, se ha predicho que al ritmo actual se alcanzará una concentración de 700 partes por millón al final del siglo XXI (IPCC, 1996).
El incremento atmosférico de dióxido de carbono ha generado dos problemas directamente relacionados. El primero, de carácter fisicoquímico, involucra no solamente al dióxido de carbono, sino que se refleja en el llamado "efecto de invernadero". A éste se atribuye el incremento en la temperatura del planeta (de más de 4 grados centígrados), ya que el dióxido de carbono (junto con el metano, el vapor de agua y otros gases en la atmósfera) tiene la propiedad de absorber luz infrarroja. Esto genera la retención de energía solar (calor) que es capaz de influir en el clima del planeta. El segundo aspecto se refiere al efecto estimulante que el incremento atmosférico de dióxido de carbono tiene en la fotosíntesis, que depende de la disponibilidad de recursos en el suelo y las relaciones de competencia entre los organismos que componen una comunidad vegetal.
A escala global, los reservorios y flujos que componen el ciclo del carbono (C) son múltiples y de diversa complejidad. (Los reservorios son depósitos de sustancias destinadas a ser usadas en un sistema; por ejemplo, el carbono fijado en los seres vivos, en los océanos, la atmósfera, etcétera. Los flujos son las tasas de intercambio de sustancias entre reservorios de un sistema, que se determinan por la diferencia entre secuestro o fijación y respiración.) Así, tanto el fitoplancton en los océanos como las plantas en los bosques y las actividades antropogénicas (por ejemplo, emisión de dióxido de carbono, la tala de bosques, etcétera) tienen un peso específico en el ciclo del carbono. Sin embargo, el aspecto central a entender es el tamaño relativo de los reservorios y la velocidad de los flujos entre éstos. El carbono se encuentra fijado en la madera de los árboles adultos de la vegetación (aproximadamente el 90 por ciento), y de éstos, el 40 por ciento se encuentra en las selvas tropicales y subtropicales (Brown y Lugo, 1982). De ahí la indudable importancia de las selvas tropicales en el ciclo global del carbono.
Este trabajo se estructura de acuerdo a los siguientes temas principales: 1) flujos y reservorios de carbono a escala global; 2) las selvas tropicales como reservorios de carbono; 3) los árboles tropicales como sumideros de carbono (un sumidero es un conducto a través del cual se sumen las cosas de un nivel de flujo a otro. Por ejemplo, la tasa de permanencia del carbono en las selvas va de decenios a cientos de años; por el contrario, el carbono en estado inorgánico -petróleo, carbón, etcétera- va de miles a millones de años. Así, un árbol en crecimiento es un sumidero); y 4) la estructura de las selvas tropicales y sus consecuencias para el ciclo global del carbono. De esta forma se sintetiza la información que permite analizar el posible papel que las selvas tropicales desempeñan en el ciclo global del carbono como consecuencia del fenómeno denominado cambio climático.
Flujos y reservorios de carbono a escala global
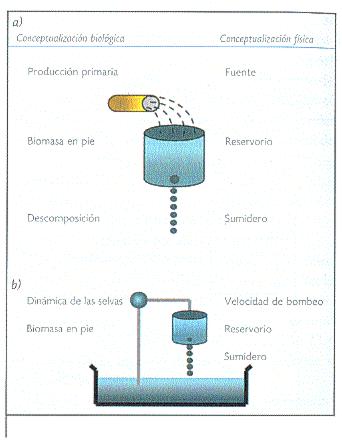
Dos principios determinan las relaciones entre los reservorios y flujos de carbono: el primero expresa que cuando un sistema (fuente-reservorio-sumidero, Figura 1a) se encuentra en balance, la tasa de suministro de la fuente es igual a la tasa de absorción del sumidero. Por ello, el tiempo de residencia del material en el reservorio está dado por el cociente entre el tamaño del reservorio y la tasa de suministro de la fuente o la tasa de absorción del sumidero. El segundo principio expresa que cuando dos reservorios de tamaño considerablemente dispar se encuentran interconectados, la tasa de transferencia entre los reservorios determinará el tamaño del reservorio más pequeño (Figura 1b).
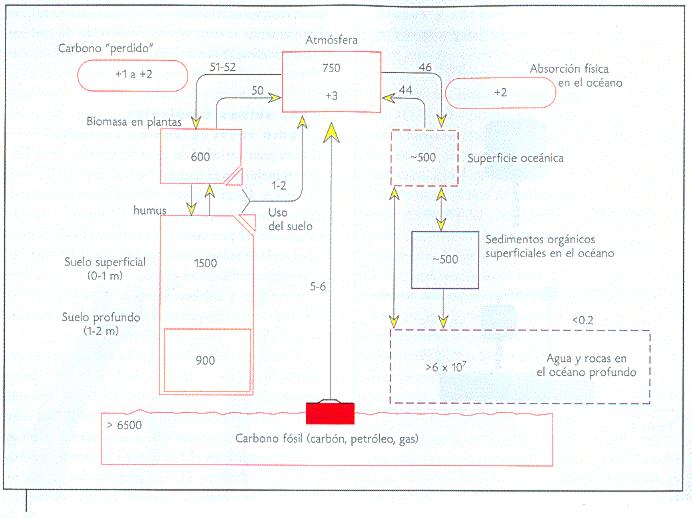
La cantidad de carbono fijado en las plantas es similar en magnitud al contenido en la atmósfera. Considerando el carbono fijado al suelo (incluyendo el humus), la biota (el conjunto de seres vivos) contiene aproximadamente tres veces más carbono fijado que el contenido actualmente en la atmósfera, y aún dos veces más que el carbono contenido en la atmósfera sumado al reservorio de carbono de la superficie oceánica (Figura 2). La tasa de recambio de carbono en la biomasa terrestre es de 14 a 19 años, y de 2 a 6 días para los organismos en el reservorio oceánico de carbono. Actualmente, el aporte antropogénico (humano) de carbono a la atmósfera (7 petagramos, es decir, 7 millones de toneladas) es de aproximadamente 1 por ciento del tamaño del reservorio atmosférico, lo cual excede en 10 por ciento el flujo natural entre la biosfera y la atmósfera.
Desde una perspectiva fisiológica, todas las plantas y algas poseen el mismo sistema enzimático bifuncional con el cual se realiza la fotosíntesis: la enzima conocida como rubisco (ribulosa bisfosfato carboxilasa). La reacción enzimática de la carboxilasa inicia la fijación de dióxido de carbono, y la reacción de la oxigenasa produce energía y pérdida de dióxido de carbono por fotorrespiración. Ambos ciclos coexisten como una reacción de competencia por dos substratos gaseosos, dióxido de carbono y oxígeno (O2). Así, si el cociente CO2/O2 cambia, se afecta la tasa de fotosíntesis neta.
|
Figura 1. Principios básicos que describen la relación entre flujos y reservorios de los elementos en los ciclos biogeoquímicos. a) Sistema fuente-reservorio-sumidero; b) Relación entre reservorios de tamaño considerablemente dispar (modificado de Walker, 1994). |
Cuando se incrementa la concentración de dióxido de carbono, se incrementa la tasa de fotosíntesis. Esto último ha sucedido de manera precipitada a partir de la revolución industrial. Sin embargo, existen una serie de factores externos (competencia, tasas de crecimiento inherentes a las especies, ritmos biológicos, etcétera) e internos que también determinan la cantidad neta de carbono fijado. Así, debido a que existe menos carbono fijado en la atmósfera que en la biota, es probable que el reservorio de carbono en la biosfera esté controlando el tamaño del reservorio de carbono atmosférico.
En otras palabras, dado el tamaño relativamente pequeño del reservorio atmosférico de carbono, en el largo plazo (al menos miles de años), lo que probablemente determina el tamaño del reservorio atmosférico es el efecto conjunto del tamaño del reservorio oceánico y la biota. Sin embargo, a una escala de tiempo corta (cientos de años), lo que codetermina el tamaño del reservorio atmosférico es la respuesta de la biota al incremento antropogénico de dióxido de carbono en la atmósfera (Körner, 2000).
Las selvas tropicales como reservorios de carbono
Una gran variedad de tipos de vegetación se pueden incluir en el término "selvas tropicales"; desde sabanas hasta bosques húmedos ecuatoriales de tierras bajas. Las selvas tropicales contienen aproximadamente 42 por ciento del carbono fijado a nivel global en la biomasa, y cerca del 11 por ciento del carbono orgánico fijado en el suelo del planeta (ver por ejemplo Olson y colaboradores, 1983; Körner, 1998). Todo esto sumado representa cerca del 50 por ciento del total del reservorio de carbono orgánico.
La mayoría del carbono en la vegetación se encuentra fijado en la madera de los árboles adultos (aproximadamente 90 por ciento), por lo que cualquier evento que modifique directamente a los árboles o indirectamente a sus procesos de regeneración tendrá un efecto inmediato en el tamaño del reservorio vegetal de carbono. Phillips (1997) propone los siguientes grupos de procesos como factores que afectan el reservorio vegetal de carbono: 1) extracción selectiva de plantas, 2) cambio en el uso del suelo, 3) fragmentación de las selvas, 4) invasión biológica, 5) cambio climático, 6) el cambio en la composición atmosférica y 7) cambios en la tasa de reposición de los árboles. De estos aspectos, el cambio en el uso del suelo, la extracción selectiva de plantas, la fragmentación de las selvas y el cambio climático son las que ejercen una mayor influencia en el papel que juegan las selvas tropicales en el ciclo global del carbono.
La extracción selectiva de plantas afecta directamente los reservorios más grandes de carbono (árboles adultos). Esto es debido a que las actividades forestales hacen uso de las especies de mayor tamaño, las cuales dominan en la estructura de la vegetación más por la biomasa acumulada que por su abundancia. Asimismo, las actividades forestales afectan principalmente las áreas que aún mantienen el 100 por ciento de la cobertura vegetal. En la actualidad, dichas áreas son extremadamente escasas. Por otro lado, la fragmentación de selvas determina que especies que naturalmente viven con bajas densidades sean especialmente vulnerables, originándose su extinción local o regional como consecuencia de la reducción de la disponibilidad de su hábitat. Asimismo, la fragmentación de selvas incrementa hasta en un 40 por ciento la mortalidad de los árboles adultos (Laurence y colaboradores, 2000).
|
Figura 2. Ciclo global del carbono. Los valores adjuntos a las flechas determinan el valor de los flujos entre reservorios en petagramos de carbono/año de flujo neto, y los valores dentro de los cuadros representan el tamaño de los reservorios, en petagramos de carbono por reservorio. Nota: el cambio en el uso del suelo y la cantidad de carbono perdido son similares (modificado de Körner, 2000). Un petagramo equivale a mil millones de toneladas. |
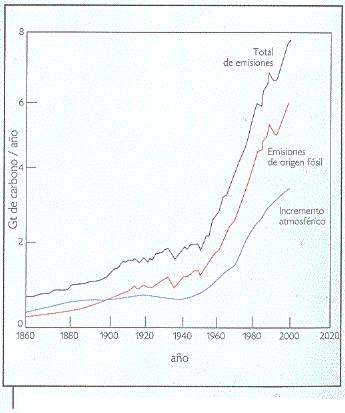
En 254 años, la concentración atmosférica de dióxido de carbono se ha elevado en un 30 por ciento (de 280 partes por millón, la concentración preindustrial, a aproximadamente 370 partes por millón en la actualidad, Figura 3). Para mitigar dicho incremento se requiere aumentar la biomasa de las selvas tropicales en 1.2 por ciento anual (aproximadamente tres billones de toneladas de carbono). Si este incremento se aplicara a una selva madura (con crecimiento de más de 100 años) se lograría un incremento anual del 11 por ciento en la ganancia neta de carbono (Körner, 2000). Un proceso de esta naturaleza estimularía fuertemente la tasa de reposición de las selvas, originando un cambio en el papel que las selvas tropicales tienen en el ciclo del carbono.
En 1994, Phillips y Gentry detectaron mediante la comparación de una extensa gama de cuadrantes de la estructura en la vegetación paleotropical y neotropical, que la tasa de reposición de árboles tropicales parecía incrementarse. Recientemente, Phillips y colaboradores (2002) reportaron un aumento en la dominancia de lianas que podría estar asociado al incremento de dióxido de carbono en la atmósfera. Si esto es cierto, la cantidad de carbono fijado por unidad de superficie no se elevará como consecuencia del efecto "fertilizante" que el incremento atmosférico de dióxido de carbono ejerce sobre la fotosíntesis. Por el contrario, tenderá a decrecer como consecuencia del incremento en la dinámica de regeneración natural de las selvas tropicales.
Los árboles tropicales como sumideros de carbono
En la vegetación, los árboles adultos son los principales reservorios de carbono. Sin embargo, en la estructura de la vegetación tropical, los árboles adultos representan solamente 1.8 por ciento del total de árboles, aunque dicho porcentaje contiene un 23.4 por ciento de la biomasa de las selvas (Laurence y colaboradores, 2000).
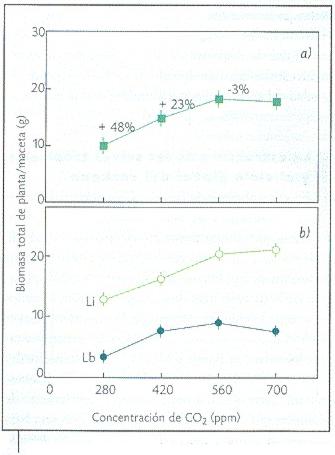
Dado el efecto estimulante que el incremento atmosférico de dióxido de carbono ejerce sobre la fotosíntesis, se podría esperar que los árboles adultos de las selvas respondan fijando una mayor cantidad de carbono. Sin embargo, existen evidencias que indican que las concentraciones actuales de carbono atmosférico se encuentran cerca del nivel de saturación. Es decir, existe un proceso límite en el sumidero de carbono que explica la respuesta asintótica temporal de la vegetación al incremento atmosférico de dióxido de carbono (Granados y Körner, 2002; Figura 4).
Es también sabido que la hoja de un árbol adulto se encuentra ligada a un reservorio de carbono de gran magnitud (ramas, tronco y raíces del árbol), de forma que la actividad del sumidero de carbono podría estar determinada por los reservorios de carbono móvil (carbohidratos no estructurales como glucosa, fructosa, etcétera) en las hojas (Körner, 2000). Sin embargo, a pesar de que en las hojas la acumulación de carbohidratos no estructurales es una respuesta al incremento atmosférico de dióxido de carbono, dicha acumulación no se encuentra necesariamente ligada a los sumideros de carbono en los árboles; más bien refleja una respuesta ligada al mesófilo de las hojas.
Otro aspecto a considerar es que la capacidad de cambiar en tamaño, masa y forma decrece con la edad del organismo en cuestión. Durante el ciclo de vida, la curva de crecimiento de cualquier planta presenta un punto de inflexión en el cual su crecimiento se hace asintótico. Esto es, una limitante espacial y evolutiva también contribuye a explicar la capacidad de fijación de carbono. Así, las primeras evidencias de que el incremento atmosférico de dióxido de carbono induce una respuesta de crecimiento que declina con la edad de la planta, fueron presentadas por Bazzaz y Miao (1983); Norby y colaboradores (1995) y Lee y Jarvis (1995). Sin embargo, dichos trabajos fueron llevados a cabo con plantas juveniles. A pesar de esto, los resultados de dichos experimentos coinciden con los reportados por Hättenschwiler y colaboradores (1997), quienes mostraron que la respuesta de árboles maduros al incremento atmosférico de dióxido de carbono depende de la edad del árbol y tiende a disminuir con la misma.
|
Figura 3. Curva comparativa del incremento de dióxido de carbono en la atmósfera. Gt: gigatonelada = 1 billón de toneladas de carbono (modificado de Joos, 1996). |
|
Figura 4. Respuesta asintótica de las plantas al incremento atmosférico de dióxido de carbono. Li = Luz intensa; Lb = Luz baja. a) Biomasa total promedio e incremento porcentual entre nivel es de dióxido de carbono; b) Biomasa total promedio por condición de luz (modificado de Granados y Körner, 2002). |
La estructura de las selvas tropicales y el ciclo global del carbono
En el sotobosque de las selvas, las plántulas crecen en condiciones de baja intensidad de luz. Esto causa que el incremento atmosférico de dióxido de carbono de lugar a una reducción en el punto de compensación de luz, la cual genera un efecto multiplicativo en la fijación neta de carbono. Es decir, a menos luz y más dióxido de carbono en la atmósfera, mayor fijación relativa de carbono. Evidencias experimentales obtenidas por Bazzaz y Miao 1983, Hättenschwiler y Körner (2000) y Granados y Körner (2002) demuestran un fuerte efecto estimulante en el crecimiento de plántulas que se desarrollan en hábitats con una baja intensidad de luz y altas concentraciones de dióxido de carbono.
De acuerdo con la hipótesis propuesta por Philips y Gentry (1994), las especies de crecimiento rápido (que fijan menos carbono por unidad de tiempo y superficie, como lianas y árboles pioneros) podrían favorecerse del incremento atmosférico de dióxido de carbono, lo que estaría influyendo en la tasa de reposición de árboles tropicales. Ésta es influida por el crecimiento agresivo de las plantas trepadoras.
Las plantas trepadoras son elementos conspicuos y característicos de cualquier ecosistema tropical, aunque se les puede encontrar en cualquier parte; aun en regiones áridas y árticas. Constituyen una forma de vida que se distingue del resto por su necesidad de soporte para desarrollarse. Esta característica implica una serie de modificaciones fisiológicas que las definen adecuadamente: 1) necesitan de un soporte para desarrollarse; por lo que tienen la capacidad de encontrar estructuras por las cuales trepar y ascender de manera eficiente hacia la luz, y pueden sobrevivir a la pérdida de su soporte mecánico; 2) poseen órganos especializados para trepar; 3) presentan en el xilema los vasos más largos y más anchos del reino vegetal, 4) se caracterizan por la asignación de un gran porcentaje de su biomasa a la superficie fotosintética y 5) las de mayor tamaño son en general leñosas, y las de menor tamaño son herbáceas.
Las plantas trepadoras tienen la capacidad de explorar una diversidad de espacios en los cuales desarrollarse, lo que les confiere ventajas competitivas a nivel del sotobosque, ya que les permite explorar ambientes de luz más heterogéneos que las plantas con modo de vida arbóreo o arbustivo. Asimismo, las lianas de mayor tamaño están igualmente representadas en diferentes tipos de suelo (desde los suelos fértiles hasta los pobres), contrariamente a la situación de los árboles más grandes, que prevalecen en suelos fértiles (Gentry, 1991). A esto se agrega el hecho de que la producción y la tasa de reposición de hojas en las plantas trepadoras es más rápida que la de muchas especies arbóreas lo que permite entender la capacidad de climatización fotosintética de una planta que se desarrolla bajo condiciones de gran heterogeneidad, como el sotobosque de las selvas.


Las plantas trepadoras pueden influir en la tasa de reposición de los árboles, ya que existe una relación negativa entre la abundancia de lianas y árboles de etapas sucesionales tardías, lo cual sugiere que las lianas reducen las tasas de supervivencia de los árboles de lento crecimiento, alterando así la fase de regeneración por claros en favor de especies de árboles de etapas de sucesión temprana (árboles pioneros). Evidencias experimentales muestran que las lianas contribuyen a la mortalidad de los árboles hospederos. Además, pueden causar daños conspicuos a los tallos y ramas de árboles jóvenes. Todos estos efectos negativos pueden, en conjunto, causar que decline el crecimiento de los árboles hospederos.
La regeneración natural de las selvas ocurre a través de la dinámica de claros, la cual inicia cuando un árbol cae y se abre un claro en el dosel, lo que da origen a un proceso de sucesión, evento clave para el establecimiento y crecimiento de los árboles tropicales. Las lianas son parte importante de dicho proceso.
Aunado a esto, en condiciones de baja intensidad de luz el incremento atmosférico de dióxido de carbono estimula el crecimiento de las plantas trepadoras. El efecto tiende a disminuir con el incremento en la concentración de dióxido de carbono (es no linear) y su influencia se acentúa en el intervalo que va de 280 partes por millón, la concentración preindustrial, a 420 partes por millón, poco menos (60 partes por millón) que la concentración actual de dióxido de carbono en la atmósfera (Figura 4). Esto demuestra que actualmente el efecto sobre la dinámica de regeneración de las selvas se encuentra en su fase más intensa.
Por otro lado, la competitividad de las plantas trepadoras se acentúa con el incremento atmosférico de dióxido de carbono. No solamente en términos de ganancia de biomasa, sino también en el incremento de la longitud del tallo y el número de internodios, lo cual confiere una mayor capacidad para explorar el entorno (Granados, 2002). Evidencias recientes indican que algunos géneros de árboles de rápido crecimiento, muchos de los cuales son árboles emergentes y de dosel, incrementan su dominancia o su densidad en las selvas prístinas de la Amazonia (Laurance y colaboradores, 2004).
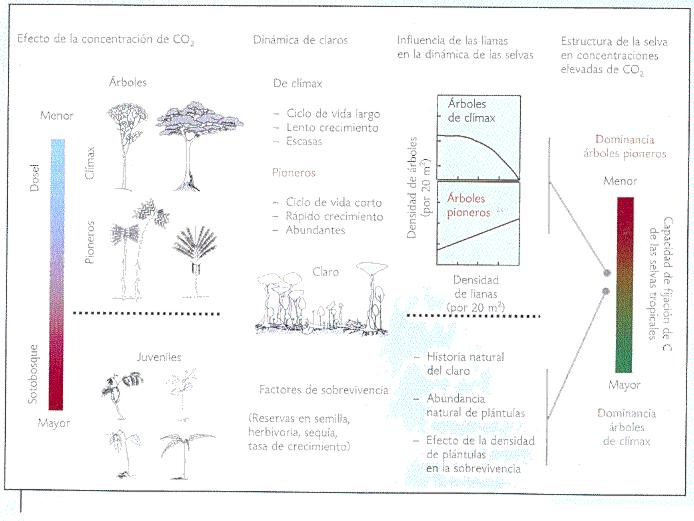
La capacidad de almacenamiento de carbono en las selvas tropicales depende del impacto que la reacción al incremento atmosférico de dióxido de carbono de las plantas trepadoras y las especies de rápido crecimiento tenga sobre la estructura de la vegetación tropical (Figura 5). Las principales implicaciones de dicho proceso son que: 1) dada la influencia de las plantas trepadoras sobre la dinámica de regeneración natural de las selvas tropicales, el efecto estimulante del dióxido de carbono sobre el crecimiento de las plantas trepadoras actualmente favorece el desarrollo de especies de sucesión temprana, en detrimento de especies de sucesión tardía; 2) las especies de sucesión temprana se caracterizan por contener menos carbono por unidad de tiempo y superficie que las especies de etapas de sucesión tardía; por lo que en el largo plazo las selvas tropicales podrían llegar a ser una fuente neta de carbono, más que un sumidero del mismo, y 3) durante los pasados 100 a 15 mil años (un periodo corto en términos evolutivos), las plantas se han desarrollado con bajas concentraciones de dióxido de carbono (280 partes por millón como máximo), por lo que se puede afirmar que están adaptadas a vivir en tales condiciones. Debido a que el incremento atmosférico de dióxido de carbono (de 280 a 370 partes por millón) se ha dado en apenas 254 años (tiempo nulo en términos evolutivos), es de esperarse un efecto de carácter no predecible hasta nuestros días sobre las relaciones bióticas (planta-planta, planta-animal, etcétera) en los ecosistemas y, por lo tanto, sobre la biodiversidad. Esto último, debido a que el incremento atmosférico de la concentración de dióxido de carbono es un factor evolutivo que favorece a algunos grupos de plantas en detrimento de otras. Además, en términos temporales, el incremento atmosférico de dióxido de carbono es un hecho inédito en la historia evolutiva del planeta. La velocidad a la que dicho cambio se ha dado y se sigue dando tiene y tendrá un efecto sobre la biodiversidad vegetal de las selvas tropicales.
|
Figura 5. Mecanismo que explica el probable impacto de las plantas trepadoras en el ciclo global del carbono (modificado de Granados, 2002). |
Un hecho concluyente para el ciclo global del carbono es que el incremento en la dinámica de regeneración de las selvas tropicales está generando un cambio en el papel que las selvas tropicales tienen en el ciclo global del carbono. Esto lleva a replantear las estrategias en la búsqueda de sumideros de carbono para mitigar el efecto que tienen las emisiones antropogénicas de dióxido de carbono sobre la biosfera y el clima del planeta.
Bibliografía
Granados, J. y C. Körner (2002), "In deep shade, elevated CO2 increases the vigor of tropical climbing plant", Global Change Biology, vol. 8, págs. 1109-1117.
Granados, J. (2002), The response of tropical liana to elevated CO2, tesis de doctorado, Universidad de Basilea, Suiza, octubre 2002.
Hättenschwiler, S., F. Miglietta, A. Raschi y C. Körner (1997), "Thirty years of in situ tree growth under elevated CO2: a model for future forest responses?", Global Change Biology, vol. 3, págs. 436-471.
Hättenschwiler, S. C. y C. Körner (2000), "Tree seedling responses to in situ CO2-enrichment differ among species and depend on understory light availability", Global Change Biology, vol. 6, págs. 213-226.
Körner, C. (2000),"Biosphere response to CO2 enrichment", Ecological applications, vol. 10, pags. 1590-1619.
Laurance, W. F., P. Delamonica, S. G. Laurance, H. L. Vasconcelos y T E. Lovejoy (2000), "Conservation: Rainforest fragmentation kills big trees", Nature, vol. 404, pág. 836.
Laurance, W. F., A. A. Oliveira, S. G. Laurance, R. Condit, H. E. M. Nascimento, A. C. Sánchez-Thorin, T E. Lovejoy, A. A, Sammya D’Angelo, J. E. Ribeiro y C. W. Dick (2004), "Pervasive alteration of tree communities in undisturbed Amazonian forests", Nature, vol. 428, págs. 171-173.
Phillips, O. L., A. H. Gentry (1994), "Increasing turnover through time in tropical forests", Science, vol. 263, págs. 954-958.
Phillips, O. L., R. Vásquez M., L. Arroyo, T R. Baker, T. Killeen, S. L. Lewis, Y. Malhi, A. Monteagudo, D. Neill, P. Núñez V., M. Alexiades, C. Cerón, A. Di Fiore, T. Erwin, A. Jardim, W. Palacios, M. Saldias, y B. Vinceti (2002), "Increasing dominance of large lianas in Amazonian forests", Nature, vol. 418, págs. 770-774.
Schnitzer, S. A., J. W. Dalling, W. P. Carson (2000), "The impact of lianas on tree regeneration in tropical forest canopy gaps: evidence for an alternative pathway of gapphase regeneration", Journal of Ecology, vol. 88, págs. 655-666.
Julián Granados nació en Oxkutzcab, Yucatán, en 1968. Cursó la licenciatura en biología en la Facultad de Biología de la Universidad Autónoma de Yucatán. Estudió el doctorado en la Universidad de Basilea, Suiza. Actualmente es investigador asociado en la Unidad de Recursos Naturales del Centro de Investigación Científica de Yucatán con el proyecto "Las selvas tropicales como sumidero de carbono". Esta dirección de correo electrónico está siendo protegida contra los robots de spam. Necesita tener JavaScript habilitado para poder verlo.