|

|
Lo malo de la endogamia El efecto perjudicial de la endogamia, también llamado “depresión endogámica”, necesariamente tuvo que ser observado por el ser humano, probablemente dentro del mismo grupo al que pertenecía, al descubrir que los hijos (progenie) de parejas emparentadas mostraban anomalías como enanismo, albinismo, hemofilia, etcétera, que se acentuaban a lo largo de las generaciones. Para evitarla debió comenzar a hacer precisamente lo contrario del apareamiento endogámico, es decir, procurar que los cruzamientos se hicieran entre individuos no emparentados.
Sin embargo, por varias razones, como la preservación del linaje, los apareamientos entre parientes se han dado a lo largo de la historia. En las dinastías faraónicas y las europeas, los matrimonios tenían lugar solamente entre miembros de la familia, a fin de mantener la “pureza” de la sangre. Caro lo pagaron al darse cuenta que en algunos individuos de las progenies se manifestaban severos defectos.
|
|
De que los genes en un locus de un individuo sean idénticos a los genes de un locus de otro individuo. Por lo tanto, la coancestría entre dos progenitores es igual a la endogamia de su progenie. De hecho, en la práctica, la endogamia de una progenie es igual a la coancestría de sus progenitores. Por ejemplo, si X y Y son los progenitores de Z, entonces rXY = FZ
Líneas endogámicas
Como ya se dijo, el apareamiento controlado dentro de varios grupos de una población, en el que los individuos dentro de cada grupo se aparean en la misma forma en cada generación, da lugar a líneas endogámicas. Esto se debe a que, siendo relativamente pequeño el número de individuos en cada línea, a lo largo de las generaciones los individuos van estando cada vez más emparentados, y son por lo tanto más endogámicos.
En teoría, en generaciones muy avanzadas las líneas son homocigóticas con F = 1, y muestran gran homogeneidad o similitud entre los individuos que las constituyen. En contraposición, las diferentes líneas muestran gran diversidad entre ellas.
La distribución estadística de un gran número de líneas endogámicas es la distribución normal. Por lo tanto en el mejoramiento de animales, si se practica selección tanto dentro de líneas como entre líneas, y se hacen cruzamientos entre los individuos seleccionados, lo menos emparentados posible, se llega a la obtención de las razas mejoradas.
En las plantas, un sistema similar es la selección recurrente; con este método se obtienen líneas y se hace selección y recombinación (cruzamiento) entre los individuos seleccionados a través de varias generaciones. Se van obteniendo líneas que, aunque son endogámicas, van siendo cada vez de mayor rendimiento, vigor, etcétera, por efecto de la selección y la recombinación. En la hibridación del maíz, aunque hay cierto tipo de selección, las líneas se usan como progenitores para obtener híbridos de mucho más alto rendimiento que las líneas progenitoras.
En el campo del mejoramiento genético del maíz por hibridación, G. H. Shull y E. M. East son considerados los descubridores de la endogamia y la heterosis en este importante cultivo, a principios del siglo pasado. Cada quien por su lado, Shull e East, descubrieron primero la endogamia y, como consecuencia de ésta, la heterosis. La endogamia, obtenida por el proceso de autofecundación, mostraba, en parcelas separadas de las líneas autofecundadas, que conforme avanzaban las generaciones, las líneas tenían plantas cada vez más uniformes, mientras que las diferencias entre las líneas eran cada vez mayores. Además, al hacer cruzamientos entre líneas, las progenies también eran tan uniformes como sus líneas progenitoras, pero de mucho mayor vigor y rendimiento que éstas. A esto fue a lo que se le llamó “vigor híbrido”, y posteriormente “heterosis”, término propuesto por Shull en una conferencia que presentó en la Universidad de Gotinga, Alemania, poco antes de estallar la Primera Guerra Mundial.
La endogamia y la heterosis, dos caras de la misma moneda
La heterosis o vigor híbrido es, entonces, la superioridad del valor genético de una progenie con respecto a sus progenitores. En el lenguaje del mejoramiento genético, nos referiremos entonces a los progenitores como las líneas, y a la progenie como el híbrido de la primera generación.
Falconer (1961) demostró que la cantidad de heterosis de las cruzas entre pares aleatorios de líneas es igual a la depresión endogámica de éstas, pero de signo contrario. Esto significa que el vigor híbrido será mayor conforme sus líneas progenitoras tengan mayor endogamia; es decir, que de esta forma la endogamia se aprovecha para generar poblaciones mejores que los progenitores.
La endogamia en el mejoramiento genético del maíz
El uso de líneas autofecundadas como progenitores del maíz híbrido no fue casual: se basó en alguna teoría desarrollada antes de que se llevara a cabo en la práctica y luego se generalizara.
La primera fue la de los caracteres cuantitativos (que se pueden medir numéricamente), en la que se propone que éstos están determinados por un grupo de genes a los que se les podían calcular sus propiedades estadísticas como la media y la varianza, así como el grado promedio de dominancia de los loci (plural de locus) involucrados. Por su parte, la teoría sobre la estructura de las líneas autofecundadas tuvo su base en lo que se llamó la “prueba temprana”. En el mejoramiento genético del maíz, a las líneas de la primera generación de autofecundación se les llama líneas S1 (S, del inglés selfing, autofecundación).
La prueba temprana consiste en cruzar una serie de líneas S1 con una “población probadora” heterogénea de plantas S0, sin selección. Por lo tanto, el rendimiento de la cruza de cada línea es el promedio de su valor genético por los valores genéticos de dicha población; a esto se le llama “prueba temprana de aptitud combinatoria general” (acg). Se espera que las líneas S1 transmitan sus capacidades a su progenie. Se infirió entonces que la obtención de líneas autofecundadas homocigóticas era el procedimiento más adecuado, ya que cumplía con un requisito indispensable en la hibridación: el mantenimiento de la pureza de las líneas en su reproducción masiva para fines comerciales de producción de híbridos.
Tipos de líneas endogámicas en el mejoramiento genético del maíz
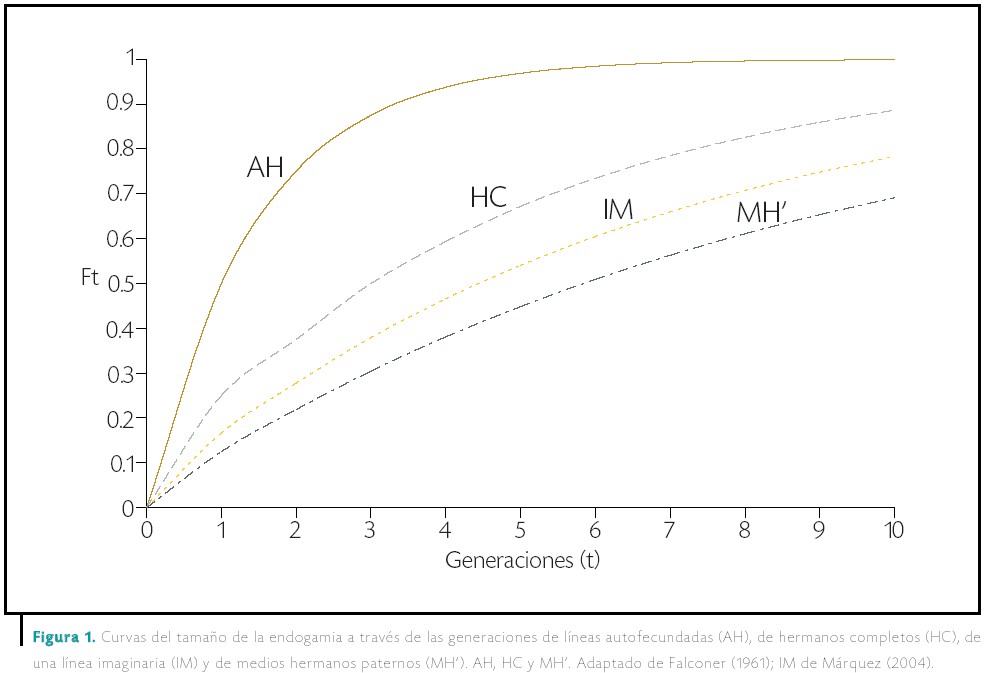
También se pueden obtener líneas homocigóticas mediante otros sistemas, como el apareamiento entre hermanos completos (por polinización fraternal) y entre medios hermanos, al cruzar una planta macho con varias plantas hembra (por polinización mesofraternal), que son los más conocidos. Estos esquemas de apareamiento que se llevan a cabo dentro de cada línea en la que se fracciona a la población se llaman “sistemas regulares de endogamia”. En la Figura 1 se muestran las curvas de endogamia de las líneas autofecundadas, fraternales, mesofraternales y de una línea “imaginaria” cuya aproximación se puede obtener en la práctica mediante un apareamiento especial (Márquez, 2005) (Figura 2).
La endogamia en las variedades sintéticas
Cuando los primeros cultivadores de maíz en Estados Unidos comenzaron a sembrar los maíces híbridos, allá en los años veinte del siglo pasado, descubrieron que las plantas en su parcela de producción comercial del híbrido eran homogéneas. Pero en la siguiente generación, sembrada con la semilla cosechada, las plantas eran muy diversas (heterogéneas) y producían considerablemente menos grano que las plantas de la primera generación.
En el Departamento de Agricultura de los Estados Unidos, S. Wright comenzó a estudiar qué pasaría en la segunda generación si en lugar de usar dos líneas progenitoras para formar un híbrido, se usaran más líneas. Encontró que al aumentar las líneas progenitoras de un híbrido, la depresión endogámica se reducía. Así, si el número de líneas progenitoras se incrementaba y se hacían entre ellas las cruzas posibles, la progenie de la mezcla de semillas conformaba lo que se denominó una “variedad sintética de primera generación”; si ésta se llevaba a la segunda generación, se lograba una reducción de la endogamia. A esta población se le llamó “variedad sintética de segunda generación”, o simplemente “sintético”. Fue hasta 1992 que Márquez dedujo la fórmula de la endogamia para sintéticos en plantas diploides, monoicas como el maíz, con varias plantas por línea, y las líneas con cualquier nivel de endogamia.
|
|
Incremento del germoplasma de maíz
En cualquier parcela en que se lleve a cabo la polinización en forma natural (polinización libre) se genera endogamia, en una magnitud que depende de cuántas mazorcas y cuántos granos por mazorca se hayan usado. En parcelas de maíces nativos (criollos) para producción comercial de grano, dichos números son muy altos; así, si una parcela se hubiera sembrado con mil mazorcas y 200 granos por mazorca, la endogamia producida en la primera generación es pequeñísima, con un valor de 1/200 000.
Sin embargo, en los métodos de mejoramiento genético las muestras son sumamente pequeñas. La selección masal es un método que usa 4 mil plantas provenientes de 200 mazorcas con 20 semillas cada una; en este caso, la endogamia en la primera generación de selección, sin considerar el número efectivo por varianza, es de 1/4 000 (Márquez, 1998). A continuación se anotan dos ejemplos para ilustrar cómo cambia la endogamia durante el mejoramiento genético del maíz.
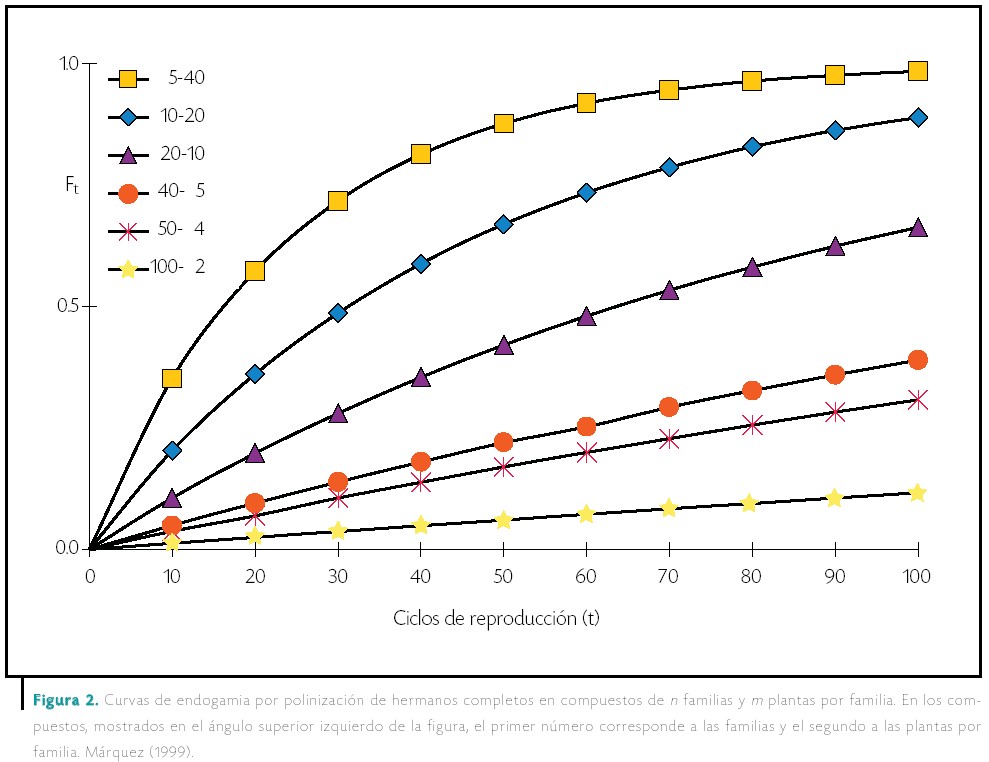
En dicho mejoramiento, el incremento de la semilla del germoplasma generalmente se hace en forma de compuestos balanceados de semillas integrados por varias (n) familias, cada familia con m plantas. El método de polinización entre las plantas que integran el compuesto puede ser fraternal o mesofraternal. En cualquier caso, lo que el fitomejorador busca es un compuesto en que predominen las familias, más que las plantas por familia. Si fuera lo contrario, habría más oportunidad de que las plantas de una familia se cruzaran entre sí y aumentara aún más la endogamia.
La forma de decidir cuántas familias y plantas usar se basa en calcular la endogamia con diferentes combinaciones de números de familias y de plantas por familia, para un total de 200 plantas, que es un número usual. Así, si se usan pocas plantas por familia para la polinización fraternal (100 familias y 2 plantas por familia), en la generación 100 de reproducción la endogamia es del orden del 10 por ciento; con polinización mesofraternal (50 familias y 4 plantas por familia), al cruzar un macho con 2 hembras, en la generación 100 la endogamia es de cerca del cero por ciento.
Es de notar que la polinización mesofraternal reduce la endogamia en mayor proporción que la polinización fraternal, porque en los medios hermanos se usa un mayor número de progenitores (más de dos) que con medios hermanos.
La endogamia en la selección masal
Quizá no haya otro aspecto más interesante de la manifestación de la endogamia que la selección recurrente. Como su nombre lo indica, ésta se lleva a cabo generación tras generación sobre una población de interés. Sin embargo, si en cada generación se seleccionara un porcentaje dado de unidades, las poblaciones resultantes serían cada vez más pequeñas, y por tanto la endogamia se iría acentuando conforme avanzaran los ciclos de selección. Después de todo, la endogamia es menos patente si las poblaciones son grandes, y sobre todo si tienen gran variación genética.
La selección masal se aplica en variedades en polinización libre. Sin entrar en los detalles de la tecnología implicada, en cualquier ciclo de selección los individuos escogidos son, obviamente, sólo una fracción del total, usualmente del 5 al 10 por ciento. Si, por ejemplo, la población inicial es de 4 mil individuos, las plantas seleccionadas serían 200 o 400. ¿Cómo entonces reconstituir el tamaño de la población si no es cruzando entre sí a los individuos seleccionados? En el primer ejemplo, si se siembran 20 semillas de cada uno de los 200 progenitores seleccionados, si todo marcha bien se tendrán nuevamente 4 mil individuos en los cuales practicar la selección. Con tal número de individuos, al cabo de 100 ciclos de selección el incremento de la endogamia es muy pequeño, de menos de 7 por ciento (Márquez, 1998).
|
|
Entonces, ¿para qué sirve la endogamia?
Por lo que hemos revisado, la endogamia sirve para producir progenies defectuosas, pero también puede servir para mejorar poblaciones. Lo segundo, desde luego, tiene que ver con la naturaleza del mejoramiento genético: el uso de poblaciones pequeñas para propósitos de selección. En este sentido, la endogamia se puede considerar como un mal necesario, pues ¿que otra forma hay para escoger, dentro de una población grande, a una muestra pequeña que tenga características deseables? No hay otra. Pero el fitomejorador, conociendo esto trata de eludir la endogamia a través de varios mecanismos; si no fuera así, no se tendrían en la actualidad las poblaciones mejoradas que han resultado de la selección en el mejoramiento de plantas, y que han sobrellevado varias decenas de ciclos de selección.
Pero la endogamia también sirve para lograr heterosis o vigor híbrido, y en este caso la endogamia es también un mal/bien necesario. Para formar híbridos vigorosos y de alto rendimiento se necesitan líneas altamente endogámicas, a veces de muy bajo rendimiento y aun con defectos que parecen insuperables, pero que al cruzarse muestran también una alta heterosis. De otra forma no se explicarían los híbridos en maíz de altos, altísimos rendimientos que se tienen en la actualidad.
|
Bibliografía
Busbice, T. H. (1969), "Inbreeding in synthetic varieties", Crop Sci. 9:601-604.
Busbice, T. H. (1970), "Predicting yield of synthetic varieties", Crop Sci. 10:260-269.
Falconer, D. S. (1967), Introduction to quantitative genetics, Oliver and Boyd, Londres, pp.85-93.
Márquez Sánchez, F. (1988), Genotecnia vegetal II, AGT Editor, México, 665 p.
Márquez-Sánchez, F. (1992), "Inbreeding and yield prediction in synthetic varieties of maize cultivars made with parental lines: I. Basic methods", Crop Sci. 32:345-349.
Márquez-Sánchez, F. (1998), "Expected inbreeding with recurrent selection in maize: I. Mass selection and modified ear-to-row selection", Crop Sci. 38:1432-1436.
Márquez-Sánchez, F. (1999), "Accumulated inbreeding in maize germplasm reproduction", Maydica 44:225-229.
Márquez-Sánchez, F. (2005), "Derivation of open-pollinated inbred lines, and their relation to Z lines for cyclic hybridización", Arnel R. Hallauer international symposium in plant breeding, CIMMYT, agosto17-22 (acepta-do para publicación).
Wright, S. (1921), "Systems of mating II. The effects of inbreeding on a genetic composition of a population", Genetics 6:124-143.
|